13.4.1. Реакции цикла Кребса относятся к третьей стадии катаболизма питательных веществ и происходят в митохондриях клетки. Эти реакции относятся к общему пути катаболизма и характерны для распада всех классов питательных веществ (белков, липидов и углеводов).
Главной функцией цикла является окисление ацетильного остатка с образованием четырёх молекул восстановленных коферментов (трёх молекул НАДН и одной молекулы ФАДН2 ), а также образование молекулы ГТФ путём субстратного фосфорилирования. Атомы углерода ацетильного остатка выделяются в виде двух молекул СО2 .
13.4.2. Цикл Кребса включает 8 последовательных стадий, обращая особое внимание на реакции дегидрирования субстратов:
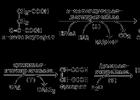
Рисунок 13.6. Реакции цикла Кребса, включая образование α-кетоглутарата
а) конденсация ацетил-КоА с оксалоацетатом , в результате которой образуется цитрат (рис.13.6, реакция 1); поэтому цикл Кребса называют также цитратным циклом . В этой реакции метильный углерод ацетильной группы взаимодействует с кетогруппой оксалоацетата; одновременно происходит расщепление тиоэфирной связи. В реакции освобождается КоА-SH, который может принять участие в окислительном декарбоксилировании следующей молекулы пирувата. Реакцию катализирует цитратсинтаза , это - регуляторный фермент, он ингибируется высокими концентрациями НАДН, сукцинил-КоА, цитрата.
б) превращение цитрата в изоцитрат через промежуточное образование цис-аконитата. Образующийся в первой реакции цикла цитрат содержит третичную гидроксильную группу и не способен окисляться в условиях клетки. Под действием фермента аконитазы идёт отщепление молекулы воды (дегидратация), а затем её присоединение (гидратация), но другим способом (рис.13.6, реакции 2-3). В результате данных превращений гидроксильная группа перемещается в положение, благоприятствующее её последующему окислению.
в) дегидрирование изоцитрата с последующим выделением молекулы СО2 (декарбоксилированием) и образованием α-кетоглутарата (рис. 13.6, реакция 4). Это - первая окислительно-восстановительная реакция в цикле Кребса, в результате которой образуется НАДН. Изоцитратдегидрогеназа , катализирующая реакцию, - регуляторный фермент, активируется АДФ. Избыток НАДН ингибирует фермент.

Рисунок 13.7. Реакции цикла Кребса, начиная с α-кетоглутарата.
г) окислительное декарбоксилирование α-кетоглутарата , катализируется мультиферментным комплексом (рис. 13.7, реакция 5), сопровождается выделением СО2 и образованием второй молекулы НАДН. Эта реакция аналогична пируватдегидрогеназной реакции. Ингибитором служит продукт реакции - сукцинил-КоА.
д) субстратное фосфорилирование на уровне сукцинил-КоА, в ходе которого энергия, освобождающаяся при гидролизе тиоэфирной связи, запасается в форме молекулы ГТФ. В отличие от окислительного фосфорилирования, этот процесс протекает без образования электрохимического потенциала митохондриальной мембраны (рис. 13.7, реакция 6).
е) дегидрирование сукцината с образованием фумарата и молекулы ФАДН2 (рис. 13.7, реакция 7). Фермент сукцинатдегидрогеназа прочно связан с внутренней мембраной митохондрии.
ж) гидратация фумарата , в результате чего в молекуле продукта реакции появляется легко окисляемая гидроксильная группа (рис. 13.7, реакция 8).
з) дегидрирование малата , приводящее к образованию оксалоацетата и третьей молекулы НАДН (рис.13.7, реакция 9). Образующийся в реакции оксалоацетат может вновь использоваться в реакции конденсации с очередной молекулой ацетил-КоА (рис. 13.6, реакция 1). Поэтому данный процесс носит циклический характер.
13.4.3. Таким образом, в результате описанных реакций подвергается полному окислению ацетильный остаток СН3 -СО- . Количество молекул ацетил-КоА, превращаемых в митохондриях в единицу времени, зависит от концентрации оксалоацетата. Основные пути увеличения концентрации оксалоацетата в митохондриях (соответствующие реакции будут рассмотрены позднее):
а) карбоксилирование пирувата - присоединение к пирувату молекулы СО2 с затратой энергии АТФ; б) дезаминирование или трансаминирование аспартата - отщепление аминогруппы с образованием на её месте кетогруппы.13.4.4. Некоторые метаболиты цикла Кребса могут использоваться для синтеза структурных блоков для построения сложных молекул. Так, оксалоацетат может превращаться в аминокислоту аспартат, а α-кетоглутарат - в аминокислоту глутамат. Сукцинил-КоА принимает участие в синтезе гема - простетической группы гемоглобина. Таким образом, реакции цикла Кребса могут участвовать как в процессах катаболизма, так и анаболизма, то есть цикл Кребса выполняет амфиболическую функцию (см. 13.1).
В отличие от многообразия макромира (мира больших и видимых невооруженным глазом существ) мир микробов характеризуется относительным однообразием. Существующие в настоящее время более 3000 различных видов бактерий, но своему внешнему виду подразделяются на 3 основные формы:
Шаровидные или эллипсовидные (кокки) размерами от 1 до 2 микрон (рис. 1.3). Кокки относятся к наиболее простой форме бактерий; они могут соединяться друг с другом, образуя диплококки (по две), тетра-кокки (по четыре) и стрептококки (цепочки); - палочковидные или цилиндрические размерами от 1 до 5 микро (рис. 1.4). Они также способны соединяться друг с другом попарно ив цепочку и дают большое разнообразие форм бактерий (диплобактерии, диплобациллы, стрептобациллы, стрептобактерии); - Извитые или спириллы размерами от 1 до 30 микрон.
Микроорганизмы-деструкторы . Ведущая роль в трансформации и минерализации органических ксенобиотиков принадлежит хемоорганотрофным (гетеротрофным) микроорганизмам, особенно бактериям, синтезирующим разнообразные ферментные системы.
Из бактерий, расщепляющих органические ксенобиотики, по частоте встречаемости, числу видов (около 30) и спектру разрушаемых соединений первое место занимают псевдомонады.
Биодеградирующая активность сообщества микроорганизмов зависит от его состава, скорости роста и обмена между видами питательными веществами и генетическим материалом. Накапливаемые метаболиты могут быть токсичны для одного компонента сообщества и могут усваиваться другими микроорганизмами, что ускоряет в совокупности процесс разложения (феномен детоксификации).
Учитывая способы получения биологических объектов - деструкторов ксенобиотиков, возможны два варианта биоочистки и биоремедиации. Первый вариант - для участков с застарелыми загрязнениями, где почти всегда обитает дикая, аборигенная микрофлора, способная их трансформировать. Такие загрязнения можно удалять in situ (по месту) без внесения биопрепаратов. При этом биодеградация лимитируется факторами окружающей среды и свойствами загрязнения, такими как содержание кислорода в среде, растворимость вещества-загрязнителя и др. Второй вариант - предварительно получают биологически активный штамм, накапливают жизнеспособные клетки, которые вносят в виде биопрепарата в загрязненную среду. Этот вариант целесообразно применять в северных регионах и при обработке мест с незастарелыми загрязнениями;
Способность микроорганизмов разрушать ксенобиотик или другой поллютант зависит от наличия в клетках генов, определяющих синтез ферментов, участвующих в деградации соединения. Конструирование рекомбинантных штаммов - деструкторов ксенобиотиков заключается в объединении нескольких генов или их блоков, ответственных за первичный метаболизм соединений. Преимущество такого объединения - генетически модифицированные микроорганизмы (ГММО) могут синтезировать различные ферментые системы, что позволяет эффективно и быстро разрушать широкий спектр химических загрязнений.
Биологическая очистка сточных вод. Принципиальные схемы очистных сооружении. Основные принципы работы, методы и сооружения аэробной и анаэробной биологической очистки сточных вод и переработки промышленных отходов.
Классификация биологических методов очистки. Биологические методы очистки применяются для очистки хозяйственно-бытовых и промышленных сточных вод (рис.2.1) от многих растворенных органических и некоторых неорганических веществ (сероводорода, сульфидов, аммиака, нитратов и др.). Процесс очистки основан на способности микроорганизмов использовать эти вещества для питания. Контактируя с органическими веществами микроорганизмы частично разрушают их, превращая в воду, диоксид углевода, нитрит-, сульфатионы и др. Органические вещества для микроорганизмов являются источником углерода. Разрушение органических веществ с помощью микроорганизмов называют биохимическим окислением.
Анаэробные микробиологические процессы осуществляются при минерализации как растворенных органических веществ, так и твердой фазы сточных вод. Анаэробные процессы протекают в замедленном темпе, идут без доступа кислорода, используются, главным образом, для сбраживания осадков. Аэробный метод очистки основан на использовании аэробных групп микроорганизмов, для жизнедеятельности которых необходим постоянный приток кислорода и температура 20-40°С.
Доступность какого-либо вещества биологическому окислению может быть оценена величиной биохимического показателя, под которым понимают отношение величин полного БПК (БПК полн) и ХПК. Биохимический показатель является параметром, необходимым для расчёта и эксплуатации промышленных биологических сооружений для очистки сточных вод. При величине биохимического показателя равном или более 0,5, вещества поддаются биохимическому окислению. Величина биохимического показателя колеблется в широких пределах для различных групп сточных вод. Промышленные сточные воды имеют низкий показатель (0,05 – 0,3), бытовые сточные воды – свыше 0,5.
Сооружения биологической очистки сточных вод. Основными сооружениями биохимической очистки служат аэротенки и вторичные отстойники.
Аэротенк представляет собой аппарат с постоянно протекающей сточной водой, во всей толще которой развиваются аэробные микроорганизмы, потребляющие субстрат, т.е. "загрязнение" этой сточной воды. Биологическая очистка сточных вод в аэротенках происходит в результате жизнедеятельности микроорганизмов активного ила. Сточная вода непрерывно перемешивается и аэрируется до насыщения кислородом воздуха. Активный ил представляет собой суспензию микроорганизмов, способную к флокуляции.
Существует также классификация аэротенков по величине "нагрузки" на активный ил: высоконагружаемые (аэротенки на неполную очистку), обычные и низконагружаемые (аэротенки продленной аэрации). Большое значение в конструкции аэротенков имеет система аэрации. Аэрационные системы предназначены для подачи и распределения кислорода или воздуха в аэротенке, а также поддержания активного ила во взвешенном состоянии.
Аэротенки-смесители (аэротенки полного смешения, рис.2, раздаточный материал) характеризуются равномерной подачей по длине сооружения исходной воды и активного ила и равномерным отводом иловой смеси. Полное смешение в них сточных вод с иловой смесью обеспечивает выравнивание концентраций ила и скоростей процесса биохимического окисления, поэтому аэротенки-смесители более приспособлены для очистки концентрированных производственных сточных вод (БПК полное до 1000 мг/л) при резких колебаниях их расхода, состава и количества загрязнений.
Аэротенки-вытеснители . В отличие от аэротенков других типов (аэротенков-смесителей и аэротенков промежуточного типа), аэротенки-вытеснители (рис.2. , раздаточный материал) представляют собой сооружения, в которых очищаемая сточная вода постепенно перемещается от места впуска к месту ее выпуска. При этом практически не происходит активного перемешивания поступающей сточной воды с ранее поступившей. Процессы, протекающие в этих сооружениях, характеризуются переменной скоростью реакции, поскольку концентрация органических загрязнений уменьшается по ходу движения воды. Аэротенки-вытеснители весьма чувствительны к изменению концентрации органических веществ в поступающей воде, особенно к залповым поступлениям со сточными водами токсических веществ, поэтому такие сооружения рекомендуется применять для очистки городских и близких по составу к бытовым промышленных сточных вод.
Аэротенки с рассредоточенным впуском (рис.2, раздаточный материал) сточной воды занимают промежуточное положение между смесителями и вытеснителями; их применяют для очистки смесей промышленных и городских сточных вод.
Аэротенки можно компоновать с отдельно стоящими вторичными отстойниками или объединять в блок при прямоугольной форме обоих сооружений в плане. Наиболее компактны комбинированные сооружения - аэротенки-отстойники. За рубежом этот тип сооружения круглой в плане формы с механическими аэраторами получил название аэроакселатора. Совмещение аэротенка с отстойником позволяет увеличить рециркуляцию иловой смеси без применения специальных насосных станций, улучшить кислородный режим в отстойнике и повысить дозу ила до 3-5 г/л, соответственно увеличив окислительную мощность сооружения.
Разновидность аэротенка-отстойника – аэроакселатор представляет собой круглое в плане сооружение. Осветленные сточные воды поступают в нижнюю часть зоны аэрации, куда пневматическим или пневмомеханическим способом подается воздух, что обеспечивает процесс биохимического окисления, а также создает циркуляционное движение жидкости в этой зоне и подсос иловой смеси из циркуляционной зоны отстойника. Из зоны аэрации иловая смесь через затопленные регулируемые переливные окна поступает в воздухоотделитель и далее в циркуляционную зону отстойника. Значительная часть иловой смеси через щель возвращается в зону аэрации, а отводимые очищенные сточные воды через слой взвешенного осадка поступают в отстойную зону.
Вторичные отстойники являются составной частью сооружений биологической очистки, располагаются в технологической схеме непосредственно после биоокислителей и служат для отделения активного ила от биологически очищенной воды, выходящей из аэротенков, или для задержания биологической пленки, поступающей с водой из биофильтров. Эффективность работы вторичных отстойников определяет конечный эффект очистки воды от взвешенных веществ. Для технологических схем биологической очистки сточных вод в аэротенках вторичные отстойники в какой-то степени определяют также объем аэрационных сооружений, зависящий от концентрации возвратного ила и степени его рециркуляции, способности отстойников эффективно разделять высококонцентрированные иловые смеси.
Иловая смесь, поступающая из аэротенков во вторичные отстойники, представляет собой гетерогенную (многофазную) систему, в которой дисперсионной средой служит биологически очищенная сточная вода, а основным компонентом дисперсной фазы являются хлопки активного ила, сформированные в виде сложной трехуровневой клеточной структуры, окруженной экзоклеточным веществом биополимерного состава.
Анаэробную очистку используют для удаления загрязнений из сточных вод, в качестве первой ступени очистки сточных вод с высокой концентрацией органических загрязнений (БПК n > 4-5 г/л), а также для переработки активного ила, других осадков и твердых отходов. Многие твердые отходы содержат целлюлозу, легче поддающуюся анаэробному разложению с образованием биогаза, чем аэробному окислению.
В ходе метаногенерации (метаногенеза) - анаэробного процесса с образованием метана - органические загрязнения конвертируются в биогаз, содержащий в основном СН 4 и С0 2 . Его можно использовать в качестве топлива. Количество выделяемого биогаза достаточно не только для компенсации энергетических затрат на анаэробное разложение, но и для использования сторонними потребителями - в котельных или нагревателях для получения пара и горячей воды, в стационарных газогенераторах для получения электроэнергии с рекуперацией тепла, в технологических процессах термосушки и сжигания осадков и др.
Биоценозы и биохимические процессы при анаэробной очистке. Формирование ценозов . Анаэробные биоценозы при очистке сточных вод могут представлять собой флокулы, биопленки и гранулы ила. Они развиваются в экосистемах с доминированием аноксигенных и анаэробных условий, в которых протекают процессы брожения, аноксигенного окисления (анаэробного дыхания) и метанообразования.
Аноксигенное окисление органических субстратов включает процессы де- нитрификации и сульфатредукции, протекающие при наличии ионов N0 3 , - N0 2 - , S0 4 2- и, как правило, при отсутствии кислорода. Эти процессы используются для удаления соединений азота и серы из сточных вод.
Основной процесс, который происходит в анаэробных условиях и используется для разложения и удаления органических загрязнений и отходов, - метаногенез. В процессе метаногенерации (часто называемым «метановым брожением») происходит разложение органических субстратов и загрязнений, обеззараживание и детоксикация стоков. В природе этот процесс протекает в различных средах с анаэробными условиями, в рубце жвачных животных, в термитниках.
Метаногенерация - сложный, многостадийный процесс, в котором исходные органические вещества последовательно превращаются в более простые с переходом значительной части углерода в метан и углекислый газ и в иловую жидкость. Метановое разложение включает три стадии анаэробного брожения (рис.5.1): гидролиза, кислотную (ацидогенную), ацетогенную и четвертую, метаногенную стадию (стадию газообразования).
В первой стадии брожения принимают участие микроорганизмы-гидролитики, обладающие целлюлолитической, протеолитической, амилолитической, липолитической, аммонифицирующей активностями. Содержащиеся в среде нитраты и сульфаты восстанавливаются бактериями-денитрификаторами и сульфатредукторами. В результате ферментативного гидролиза целлюлоза и гемицеллюлозы, белки, жиры и другие компоненты гидролизуются с образованием жирных кислот, глицерина, пептидов, аминокислот, моно- и дисахаров и в небольшом количестве уксусной кислоты, метанола, аммиака, водорода. В гидролизе участвуют бактерии pp. Clostridium, Bacillus, а также Bacteroides, Butyrivibrio, Cellobacterium, Eubacterium, Bifidobacterium, Lactobacillus, Selenomonas. На ацидогенной стадии протекают различные пилы брожения: спиртовое, маслянокислое, ацетоно-бутиловое, пропионовое и другие, в ходе которых бактерии-ацидогены сбраживают образующиеся продукты гидролиза, например глюкозу, до органических кислот:
Потребляя моно- и олигосахариды, аминокислоты и другие промежуточные продукты гидролиза, эти бактерии тем самым предотвращают ингибированис продуктами гидролиза гидролитических ферментов, участвующих в первой фазе брожения.
В результате расщепления на первых двух стадиях 70-80% из образующихся органических продуктов составляют высшие жирные кислоты, до 20% - ацетат и 3-5% - водород. Среди других продуктов - изомасляная, фенилуксусная, бензойная, индолилбензойная кислоты, NH 4 + , H,S, бутанол, пропанол, С0 2 и др.
На ацетогенной стадии брожения гетероацетогенные бактерии (ацетогены) pp. Clostridium, Syntrophus и других переводят органические кислоты, например пропионовую и масляную,прочие продукты ацидогенеза в уксусную кислоту:

Основную роль в метановом разложении играет заключительная стадия, выполняемая строгими анаэробами - метанообразуюшими бактериями. Они более чувствительны к условиям среды. Время генерации клеток метаногенов составляет несколько суток. Их активность максимальна при рН среды от 6,8 до 7,5. При более низких и высоких значениях рН развитие метаногенов замедляется либо прекращается совсем.
Продукт реакций метаногенной стадии - СН 4 . Образование его возможно двумя путями. Метаногенные бактерии-литотрофы (pp. Methanococcus, Methanobacterium, Methanospirillum, Methanomicrobium, Methanogenium, Methanothermus, Methanobrevibacter) потребляют в качестве субстрата Н 2 и С0 2 , а также СО и формиат:
С0 2 + 4Н 2 → СН 4 + 2Н 2 0
4НСООН → СН 4 + ЗС0 2 + 2Н,0
4СО + 2Н 2 0 → СН 4 + ЗС0 2
Микроорганизмы-ацетотрофы (pp. Methanosarcina, Melhanosaeta, Methano- planus) используют ацетат, метанол, метиламин:
CH 3 СООН → CH 4 + CО 2
4СН 3 ОН → ЗСН 4 + С0 2 + 2Н 2 0
4CH 3 NH 2 + 2Н 2 0 → ЗСН 4 + 4NH 3 + СО
Вследствие разрушения органических кислот рН среды повышается, реакция среды становится шелочной, поэтому метаногенную стадию иногда называют «щелочным брожением».
При разложении уксусной кислоты образуется 70-75% метана, а остальные 25-30% - в результате синтеза из углекислоты и водорода и протекания других реакций. Соотношение конечных продуктов в процессе метанового брожения зависит от состава среды, условий ферментации и присутствующей микрофлоры.
Большим стимулом к развитию многих из современных методов анаэробной очистки послужило обнаружение в середине 1970-х гг. способности микроорганизмов, входящих в состав метаногенного сообщества, образовывать агрегаты - гранулы (пеллеты) при росте в анаэробном реакторе в условиях восходящего потока (рис. 5,2 раздаточный материал).
Особую роль в формировании и функционировании гранул выполняют метаногенные бактерии Methanosaeta concilii (Methanothrix soehngenii), а также Methanosarcina spp. Бактерии p. Methanosaeta образуют хворосто- и клубкообразные структуры (рис. 5.3), внутри которых группируются микроколонии бактерий Methanosarcina (рис. 5.4). Благодаря этому формируются агрегаты в виде плотных легко оседающих гранул размером 1-5 мм.
К сооружениям традиционных конструкций относятся септитенки, осветлители-перегниватели, контактные реакторы, анаэробные лагуны, метантенки, анаэробные биофильтры с восходящим потоком жидкости (см.раздаточный материал, рис. 3.5).
Септитенк (септик) представляет собой аппарат, состоящий из двух частей: отстойной и септической (рис. 6.1). В первой части происходит осветление воды вследствие движения ее с малой скоростью, а во второй части, расположенной под первой, - перегнивание осадка при хранении его в течение 6-12 мес. Отстойная и септическая части септика не разделены между собой. Продолжительность нахождения воды в септике - 3-4 сут. Септики применяются, если количество сточных вод не превышает 25 м 3 /сут.
Септитенки часто используются для сбраживания активного ила вторичных отстойников, осадков первичных отстойников и пены с целью накопления осадка, уменьшения его объема, дурного запаха и количества патогенной микрофлоры. Септики являются наиболее распространенными очистными сооружениями для индивидуальных домохозяйств, поскольку могут работать автономно и не нуждаются в электропитании.
Осветлители-перегниватели, которые можно рассматривать как разновидность септитенка, применяют на станциях очистки сточных вод с пропускной способностью до 30000 м 5 /сут. На рис. .2 приведена конструкция осветлителя - перегнивателя, выполненного в виде комбинированного сооружения, состоящего из осветлителя, концентрически располагаемого внутри перегнивателя.
Метод анаэробной очистки в контактном реакторе был одним из первых, широко применяемых в промышленности с начала 1930-х гг., в частности, для обработки стоков сахарных, спиртовых и дрожжевых производств. По сравнению с септитенком контактный реактор намного производительнее, поскольку в нем предусматривается перемешивание среды с анаэробным илом и поддерживается более высокая концентрация ила за счет возврата его части из вторичного отстойника (см.раздаточный материал к лекции3, рис. 3.5), т. е. аналогично тому, как это реализуется в аэротенке с вторичным отстойником. Для повышения эффективности разделения иловая жидкость перед вторичным отстойником может дополнительно подвергаться дегазации (в отдельной емкости) или охлаждению. При дегазации газ удаляют механическим (гидравлическим) способом или действием вакуума. Охлаждение приводит к замедлению процессов метанообразования и, как следствие, образования новых пузырьков, что улучшает седиментационные свойства анаэробного ила.
Традиционными и наиболее распространенными аппаратами для проведения анаэробного разложения являются метантенки. Они используются для сбраживания стоков с высокой концентрацией загрязнений и разложения органических отходов, в частности, активного ила очистных сооружений.
Метантенки работают с обогревом, как правило, в периодическом режиме загрузки отходов или сточных вод, с постоянным отбором биогаза и выгрузкой твердого осадка по мере завершения процесса. Их изготавливают из стали, бетона, пластмасс, кирпича; они различаются формой резервуара, количеством камер сбраживания, способом загрузки, выгрузки субстрата, способами обогрева и перемешивания.
Метантенки большого объема изготавливают в виде вертикальных резервуаров цилиндрической или эллипсоидной формы с принудительным перемешиванием сбраживаемой массы, они рассчитаны на избыточное давление газа до 5 кПа. Небольшие биогазовые установки могут представлять собой цилиндрические горизонтальные или вертикальные биореакторы с механическим перемешиванием, частично или полностью заглубленные в грунт для снижения тепло- потерь. Конструкция биореакторов должна обеспечивать возможность полного опорожнения резервуара, поэтому днише часто выполняется скошенным, полусферическим или в форме конуса.
Метантенки с неподвижным незатопленным перекрытием имеют недостаток, свойственный сооружениям с жесткими перекрытиями - непостоянство давления внутри реактора. При выгрузке осадка внутри метантенка может образоваться вакуум, а при загрузке повыситься давление. Это приводит к разрушению конструкций, образованию трещин.
Преимущества метантенка с плавающим перекрытием: 1) взрывобезопасность, так как независимо от наполнения метантенка в нем поддерживается положительное давление газа, что исключает возможное попадание воздуха внутрь сооружения; 2)по положению плавающего перекрытия можно осуществить дозировку загрузки и выгрузки; 3)облегчается борьба с образованием корки.
Роль перемешивания и температурного режима в метатенках.Метантенки всех типов могут работать в мезофильном (20-45 °С, обычно 30-35 °С) и термофильном (50-60 °С) температурных режимах. Режим сбраживания выбирают с учетом методов последующей обработки и утилизации осадков, а также санитарных требований. Мезофильный режим используется чаше, поскольку он является менее энергозатратным и более экономически выгодным, допускает существование большего числа видов микроорганизмов и поэтому более стабилен, менее чувствителен к изменениям условий окружающей среды; осадки в этом режиме после переработки обезвоживаются лучше по сравнению с термофильным процессом. Однако при термофильном режиме скорость распада органических соединений выше (примерно в 2 раза) и выше степень их распада, достигается практически полная дегельминтизация осадков, что важно, если осадки используются в качестве рекультиванта или удобрения для почвы. Продолжительность сбраживания при мезофильном режиме 20-30 сут., при термофильном - около 10 сут. Теплотворная способность газа при термофильном сбраживании на 5% ниже, чем при мезофильном.
Для более полного проведения процесса метаногенерации необходимо тщательное перемешивание содержимого метантенка, чтобы обеспечить равномерное распределение содержимого реактора, необходимые условия массо- и теплообмена, свести к минимуму слипание, образование пены и корки, формирование придонного осадка, удалить газы. Для перемешивания в метантенке используют механические мешалки, циркуляционные насосы, гидроэлеваторы или комбинацию этих систем.
Оптимальная концентрация взвешенных веществ в метантенке, при которой наблюдается высокая интенсивность метанообразования, находится в интервале 2-10%. При концентрации твердых частиц выше 10-12% перемешивание среды затрудняется, и это приводит к снижению газовыделения. В таких случаях применяются специ&чьные конструкции биореакторов, обеспечивающие необходимый уровень перемешивания.
Метанообразование протекает с максимальной скоростью при рН от 6 до 8. При понижении рН ниже 5,5 (в случае «закисания» метантенка) активность метаногенных бактерий прекращается. Как правило, рН не регулируют вследствие высокой буферной емкости среды. Но при закислении среды наилучшим нейтрализующим средством является раствор NaHC0 3 .
Процесс метаногенеза замедляется в присутствии различных детергентов (при их концентрации около 15 мг/л), антибиотиков и других веществ. Из анионогенных ПАВ сравнительно полно распадаются и слабо тормозят процесс сбраживания алкилсульфаты, хлорный сульфанол; трудно поддаются распаду и сильно тормозят сбраживание сульфанолы.
Анаэробные реакторы устойчивы к длительным перерывам в подаче сточной воды, изменениям химического состава поступающих стоков, что позволяет эффективно использовать их для очистки стоков сезонных производств, а также в режимах малых нагрузок. В случае снижения метаногенной активности для ее восстановления можно снижать скорость подачи субстрата, подщелачивать среду химическими веществами, разбавлять стоки водой, удалять токсические соединения путем предварительной обработки стоков.
Бактериальное выщелачивание химических элементов из руд, концентратов и горных пород, обогащение руд, биосорбция металлов из растворов. Удаление серы из нефти и угля. Повышение нефтеотдачи. Удаление метана из угольных пластов. Подавление биокорозии нефтепродуктов.
Исследования по бактериальному окислению железа и выщелачиванию металлов начаты после выделения в 50-х годах ХХ века из дренажных кислых вод угольной шахты микроорганизмов, способных принимать участие в окислении двухвалентного железа до трехвалентного – бактерий Acidithiobaccilus ferrooxidans (ранее называвшихся Thiobaccilus ferrooxidans). Бактерии, участвующие в выщелачивании металлов, по типу питания относятся к хемоавтотрофным, для получения энергии катализирующие химические окислительно-восстановительные реакции и ассимилирующие углекислый газ для конструктивного обмена клеток, т.е. питающихся автономно, без использования органики.
Кучное биовыщелачивание сульфидных руд .
В последние годы для подготовки упорного сырья к цианированию начали использовать чановое бактериальное выщелачивание концентратов или руд. В мире уже действует более десятка промышленных предприятий, практикующих эту технологию, но капитальные затраты для такой технологии весьма высоки, поэтому для малых и средних месторождений не оправданы.
Использование строго ацидофильных бактерий предполагает, что величина рН пульпы или раствора 2 и ниже. Если для выщелачивания используют бактерий A. ferrooxidans, то процесс биологического окисления минералов может идти двумя путями: эти бактерии не только окисляют серные соединения, но и способны для получения энергии окислять закисные формы железа до окисных. Время обработки зависит от состава сульфидной руды, формы и распределения металла в руде и количества доступной для микроорганизмов серы. Существует также ряд более узких проблем, например, токсичность высоких концентраций добываемых тяжелых ценных металлов для некоторых видов или штаммов выщелачивающих микроорганизмов.
Таким образом, одним из подходов к совершенствованию и развитию технологии и методов биовыщелачивания является подбор бактерий и архей, устойчивых к токсичности металлов. Другими критериями подбора культур являются: простота работы с ними в промышленных условиях, активность в окислении минеральных соединений, отношение к рН, температуре, режиму аэрации и возможность стимулировать их активность внесением дополнительных веществ.
В настоящее время известен ряд родов (групп, подразделяемых по свойствам и систематическому положению) бактерий и архей (два надцарства микроорганизмов), представителей которых способны к выщелачиванию металлов путем окисления сульфидов: Acidothiobacillus, Halothiobacillus, Thiobacillus, Leptospirillum, Acidiphilium, Sulfobacillus, Ferroplasma, Sulfolobus, Metallosphaera и Acidianus. Таким образом развитие технологий биовыщелачивания может опираться как на внесение изменений в организацию процесса (оптимизация аэрации, температурного режима, предподготовки минерального сырья и т.п.), так и в подбор новых микробных культур – с более высокой активностью или проще наращиваемой биомассой, или с более широким спектром рН, температуры и т.п. Традиционное выщелачивание кислыми растворами привело к тому, что поиск новых культур микроорганизмов сосредоточен именно на ацидофильных и суперацидофильных организмах.
Углеводы, белки и жиры в организме гидролизуются, а образующиеся при этом продукты гидролиза – моносахариды, аминокислоты, жирные кислоты и глицерин подвергаются превращениям, в ходе которых часть из них окисляется до углекислого газа и воды, являющимися продуктами окисления углерода и водорода. Если бы система, в которой каждый из продуктов гидролиза биополимеров, представляющий собой субстрат для последующего окисления, имел бы свой метаболический путь, то такая система была бы очень громоздкой и ненадежной. Однако, Природа решила задачу унификации метаболических путей, организовав катаболические процессы таким образом, что на промежуточных этапах этих процессов образуется минимальное число одних и тех же метаболитов, которые получаются при окислении разных веществ. И, действительно, как видно из схемы, большинство субстратов окисления превращаются в пировиноградную кислоту – пируват (С 3), а затем в ацетил-КоА (С 2), причем последний может образовываться и при окислении пирувата. Ацетил-КоА полностью окисляется в цикле трикарбоновых кислот (ЦТК – он же цикл Кребса или цитратный цикл). Цикл Кребса является общим путем катаболизма для углеводов, белков и жиров. Энергия, выделяющаяся в ходе катаболических реакций, частично рассеивается виде теплоты, большая же ее часть расходуется в анаболических реакциях. Передача энергии осуществляется с помощью интермедиаторов, основной из них – АТФ. Эндергонические процессы – это синтез аденозинтрифосфата (АТФ) из аденозиндифосфата (АДФ) и неорганического фосфата, а также синтез и других веществ с макроэргическими связями. Этот процесс протекает благодаря сопряжению энергии с катаболическими реакциями. Экзергонический процесс – это гидролиз АТФ, а также других трифосфатов. Гидролиз поставляет необходимую энергию для биосинтеза.
Ниже приведена схема сопряжения анаболических и катаболических процессов:
S 1 окисленный субстрат, ΔG < 0АДФ + фосфат АТФ + Н 2 О, ΔG < 0
Сопряжение
АТФ + Н 2 О → АДФ + фосфат, ΔG < 0S 2 продукт биосинтеза, ΔG > 0
Большая часть АТФ в организме образуется в результате окислительного фосфорилирования , которое происходит в цепи передачи электронов (ЦПЭ). Основными субстратами этого процесса являются НАД*Н и ФАД*Н 2 , образующиеся преимущественно в ЦТК, поэтому одной из основных задач катаболизма является синтез АТФ – своеобразного аккумулятора энергии, необходимого для последующих реакций анаболизма. Большинство биосинтезов носят восстановительный характер, так как продукты биосинтеза являются менее окисленными по сравнению с исходными веществами. Роль восстановителя в таких процессах играет НАД*Н. Таким образом, ключевая роль в метаболизме принадлежит ограниченному числу соединений. Это пируват и ацетил-КоА, вещества которыми заканчиваются специфические пути катаболизма; АТФ, продукты гидролиза, к которым поступает энергия для анаболических процессов; НАД*Н и ФАД*Н 2 – коферменты, при окислении которых образуется основная часть АТФ в организме.
Катаболизм углеводов
Процессы обмена углеводов у человека начинаются в ротовой полости, так как в состав слюны входит фермент амилаза, который способен расщеплять крахмал и гликоген до дисахарида – мальтозы, которая ферментом мальтазой расщепляет последнюю до глюкозы. Поступление глюкозы в клетки различных органов зависит от гормона инсулина, который регулирует скорость переноса глюкозы через мембраны клеток. переносчиками – белками.
Обмен глюкозы в клетке начинается с ее фосфорилирования:
Глюкоза + АТФ глюкозо-6-фосфат + АДФ


АТФ → + АДФ
В отличие от свободной глюкозы, глюклзо-6-фосфат не способен проходить через клеточные мембраны, поэтому фосфорилированная глюкоза как бы «запирается» в клетке, и там запасается в форме гликогена – животного крахмала, который синтезируется из молекул глюкозо-6-фосфата.
Катаболизм глюкозы в клетке может идти по трем основным направлениям, которые различаются по способу изменения углеродного скелета молекулы:
1. Дихотомический путь, при котором происходит расщепление связи С-С между третьим и четвертым атомами углерода, и из одной молекулы гексозы получаются две триозы (С 6 →2С 3).
2. Апотомический путь (пентозофосфатный), при котором гексоза превращается в пентозу (С 6 → С 5) в результате окисления и отщепления одного (первого) углеродного атома.
3.Глюкуроновый путь, когда происходит окисление и отщепление шестого углеродного атома
Главным путем распада глюкозы, ведущим к высвобождению энергии является дихотомический путь, а в этом пути, в свою очередь, окислить глюкозу и получить ее энергию можно двумя способами:
1.Независимый анаэробный распад глюкозы до молочной кислоты – гликолиз.
глюкоза →2-лактат + 134 кДж
Часть этой энергии расходуется на образование двух молекул АТФ, а остальная рассеивается в виде теплоты.
2.Аэробный (кислородзависимый) распад глюкозы до углекислого газа и воды
Это процесс обратный фотосинтезу:
С 6 Н 12 О 6 + 6О 2 ↔ 6СО 2 + 6Н 2 О + 2850 кДж
60% этой энергии запасается в виде макроэргических связей АТФ, то есть в биологически доступной форме. Как видно из приведенных уравнений, аэробный путь, несомненно, более выгоден по сравнению с гликолизом, так как в нем из одинакового количества глюкозы образуется в двадцать раз больше АТФ. Аэробный распад осуществляется большинством тканей организма за исключением эритроцитов. Для злокачественных клеток основной путь получения энергии – гликолиз. Мышцы используют гликолиз в случае больших нагрузок, когда затруднен доступ кислорода и тогда в натруженных мышцах образуется молочная кислота.
Цепь реакции гликолиза глюкозы включает в себя одиннадцать реакций, из которых первые десять - общие с аэробным распадом, а одиннадцатая – это синтез молочной кислоты из пировиноградной кислоты (ПВК) с помощью НАД*Н. Рассмотрим последовательно реакции при аэробном распаде глюкозы:
1 реакция – это фосфорилирование глюкозы, ее активация.
2 реакция – это изомеризация, глюкозо-6-фосфат превращается в фруктозо-6-фосфат.
3 реакция - фруктозо-6-фосфат фосфорилируется до фруктозо-1,6-дифосфата.
Первые три реакции представляют собой так называемую подготовительную стадию, на этом этапе еще идет затрата энергии АТФ на реакции фосфорилирования:
1Глюкозо-6-фосфат
2- изомеризация
|
2-фосфоглицерат общий путь
9 Н 2 О АТФ
Следующий этап – это реакции гликолетической оксиредуктации , в которых идет распад шестиуглеродного скелета на два трехуглеродных и окисление их до пирувата.
4 реакция - фруктозо-1,6-дифосфат в своей открытой ациклической форме распадается с помощью фермента альдолазы на два трехуглеродных фрагмента: глицеральдегидфосфат и диоксиацетонфосфат.
5 реакция - изомеризация, превращение диоксиацетонфосфата в глицеральдегидфосфат.
Дальнейший катаболизм происходит только через глицеральдегидфосфат, две молекулы которого в 6-ой реакции окисляются НАД + в 1,3-дифосфоглицерат, а выделяющаяся при этом энергия запасается в виде АТФ. В данном случае окисление альдегида приводит к ангидриду органической и фосфорной кислоты. Две молекулы 1,3-дифосфоглицерата превращаются в процессе гидролиза в 3-фосфоглицерат, а далее, в 8-ой реакции происходит перенос фосфатной группы из положения 3 в положение 2.
9 реакция- отщепление воды с получением фосфоенолпирувата, а затем происходит кето-енольное превращение, сопряженное с гидролизом, когда от диоксиацетонфосфата отщепляется одна молекула фосфорной кислоты и енольная форма превращается в кетоформу.
КАТАБОЛИЗМ ЛИПИДОВ
У высших животных и человека липиды поступают в желудок, и выходят из него почти не затронутые кислой средой. В щелочной среде тонкого кишечника липиды гидролизуются под действием липаз. Гидролизованные липиды всасываются в кровь и переносятся в различные органы для дальнейшего метаболизма.
В кровь поступают сквозь стенку кишечника глицерин, ЖК, моно- и диглицериды. В крови ЖК снова этерифицируются глицерином, который связан с белками крови и переносится в жировую ткань или печень, где откладывается. В печени идёт гидролиз с образованием ЖК, которые окисляются до СО 2 и Н 2 О. При окислении высвобождается большое количество энергии.
Процесс окисления ЖК включает много стадий. ЖК разрушается (синтезируется) до фрагментов С-С (природные ЖК состоят из чётного числа атомов углерода). При катаболизме, ЖК сначала превращаются в тиоэфиры с коферментом А, с выделением АТФ, затем окисляются в ненасыщенные кислоты, окислителем служит ФАД.
С 15 Н 31 СООН – пальмитиновая кислота
О HSКоА О ФАД
СН 3 (СН 2) 12 СН 2 СН 2 С ОН СН 3 (СН 2) 2 СН 2 СН 2 С SКоА
СН 2 (СН 2) 12 СН=СНС SКоА
Начинается путь катаболизма белков с гидролиза (протеолиза) под действием ферментов протеазы и пептидазы.
Гидролиз белков начинается в желудке под действием фермента пепсина, этому способствует кислая среда желудочного сока рН=1-2 возникает благодаря выделению желудочных клеток соляной кислоты.
В тонком кишечнике при рН=7,8-8,4 , распад белков катализируется ферментами поджелудочной железы трипсином и химитрипсином.
АК – продукт гидролиза белков, поступающие из ЖКТ, являются важным фондом пополнения аминокислотного запаса клеток и тканей. Ограниченное поступление из вне даже одной из незаменимых АК вызывает резкий распад собственных белков тканей, АК используются в синтезе собственных белков, нуклеотидов, порфиринов и т. д.
В сутки взрослому человеку необходимо 100 г белка. Белки могут быть полноценными – в наличии все незаменимые АК и неполноценными – в наличии не все незаменимые АК. За сутки распадается и синтезируется 400 г белка. За 35 дней обновляются все белки.
О состоянии белкового обмена можно судить по азотистому балансу. Поскольку белки органов отличаются строгой видовой и тканевой специфичностью, живой организм обладает способностью использовать вводимый белок только в гидролизованном состоянии.
Всасывание АК через мембрану тонкого кишечника происходит под действием глутатиона. АК поступают в кровь воротной вены, затем в печень, где подвергаются ряду превращений.
Отправить свою хорошую работу в базу знаний просто. Используйте форму, расположенную ниже
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
Курсовая работа
на тему:
« Биохимические пути в исследовании механизмов психических и нервных болезней »
ВВЕДЕНИЕ
Патологические состояния центральной нервной системы многочисленны, многообразны и чрезвычайно сложны по механизму возникновения и развития. В этой работе будут показаны только пути, на которых ученые-биохимики добились некоторых успехов в познании отдельных элементов патологических процессов, лежащих в основе ряда болезней и болезненных состояний центральной нервной системы.
1. ХИМИЧЕСКИЕ ФАКТОРЫ ВНУТРЕННЕГО ПОДКРЕПЛЕНИЯ ПРИ НАРКОМАНИЯХ
У примитивно устроенных организмов цепочка, реакций в центральной нервной системе, начинающаяся поя действием той или иной мотивации, например голода, инстинкта продолжения рода и т.п., имеет минимальное число промежуточных звеньев и завершается непосредственным достижением или недостижением цели. Достижение конечной цели является простейшим подкреплением, вознаграждающим фактором. У относительно высокоразвитых организмов процесс достижения цели может быть разделен на большее число этапов. Завершение промежуточного этапа не вознаграждается конечным результатом и, как установлено, существует система так называемого внутреннего подкрепления. Например, выполняя определенную работу, современный человек, как правило, не получает после ее завершения пищевого вознаграждения, но получает деньги, испытывая при этом определенное удовлетворение и уверенность в возможности с помощью денег реализовать цель - приобретение и потребление пищи. В центральной нервной системе чувство удовлетворения может быть обеспечено с помощью ряда гуморальных факторов, к которым в первую очередь относятся нейропептиды - некоторые из опиоидов, нейротензин и др.
Простейший эксперимент, позволяющий в опытах на животных выявить эти гуморальные факторы, состоит в предоставлении им возможности выбора веществ, вызывающих приятные ощущения при самовведении в желудочек мозга. Белая крыса, например, с вживленными в мозг канюлями, имеющая возможность нажатием той или иной педали инъецировать себе раствор одного из испытуемых веществ, довольно быстро переходит от беспорядочного нажатия разных педалей к заведомо предпочтительному самовведению довольно узкого круга веществ. Существуют и более сложные формы такого рода экспериментов.
В результате, к категории предпочитаемых веществ - вероятных внутренних факторов подкрепления - сейчас относят некоторые из опиоидных нейропептидов - р-эидорфин и эике-фалины , а также нейротензин. Одновременно выявляются и нейропептиды, обладающие противоположным действием: вазопрессин и, по-видимому, меланостатин и тиролиберин.
Вводя себе извне нейропептиды вознаграждения, животное обходится без нормального механизма, необходимого в естественных условиях для обеспечения пути к цели, завершение которого вознаграждается образованием внутреннего химического сигнала - фактора внутреннего удовлетворения.
Многие исследователи рассматривают эти эксперименты как модель наркомании. Существо наркомании состоит, с этой точки зрения, в подмене внешним химическим агентом естественного внутреннего химического вознаграждения. Такая подмена при доступности химического эквивалента внутреннего фактора не требует целенаправленного труда и ряда других процессов для того, чтобы достичь состояния удовлетворения, наслаждения и т.п. Экзогенные опиаты - морфин и его аналоги - являются эквивалентами внутренних опиоидов, взаимодействуя с теми же классами рецепторов головного мозга, что и опиоидные нейропептиды. Характерно, что наиболее апробированным средством снятия абстинентных состояний у наркоманов является специфический блокатор опиатных рецепторов - налоксон, а также его аналоги.
Механизмы наркоманий, вызванных опиатами, находят, таким образом, истолкование, которое можно отнести к категории достаточно обоснованной гипотезы. Сложнее обстоит дело с рядом других наркотиков - кокаином, каниабиноидами, ЛСД, мецкалинами и др. В отношении некоторых из них рассматриваются гипотезы, аналогичные изложенной выше. В частности, катехо ламины, особенно норадреналин, и серотонин, в определенных зонах мозга участвуют в процессах внутреннего подкрепления. Об этом свидетельствует наличие зон, расположенных по ходу катехоламинергических и серотонинергических путей, раздражение которых вызывает ощущения удовлетворения, удовольствия и т.п. Многие из упомянутых выше наркотинов известны как агенты, вмешивающиеся в катехоламинергическую и серотонинергическую нейротрансмиссию. Поэтому, хотя и с меньшей степенью доказательности, чем в случае опиатных наркотиков, гипотеза о наркомании, как подмене факторов внутреннего подкрепления, правомочна и для многих неопиатных соединений. Следует также иметь в виду, что механизмы и химические факторы внутреннего подкрепления известны лишь частично.
В рамки изложенной гипотезы пытались заключить и данные о механизмах алкоголизма. Прослеживаются сложные связи между развитием алкоголизма, уровнями опиоидных пептидов, непептидных факторов, подобных опиатам, катехоламинов, серотонина и других гуморальных регуляторов. Сложность этих связей такова, что, не исключая роли факторов внутреннего подкрепления, целесообразно рассмотреть ряд данных о механизмах алкоголизма особо.
2. АЦЕТАЛЬДЕГИД, НЕПЕПТИДНЫЕ И ПЕПТИДНЫЕ ОПИОИДЫ И АЛКОГОЛИЗМ
Первые исследования биохимических механизмов алкоголизма привели к установлению трех важных фактов. Во-первых, этанол является мембранотрогтным агентом и может в концентрациях, вызывающих опьянение, менять состояние рецепторов и многих энзимов, инкорпорированных в мембрану. Во-вторых, широко представленный в организме, особенно в печени, фермент - алкогольдегидрогеназа, участвующая в метаболизме многих регуляторных соединений, вступает в контакт с поступающим извне этанолом и как бы отвлекается от ряда нормальных функций, что, в свою очередь, ведет к отклонениям в синтезе ряда регуляторов. В-третьих, наконец, алкогольдегидрогеназа быстро превращает часть поступающего этанола в ацетальдегид; последний может быть источником образования ряда биоактивных факторов и, кроме того, его прямое действие на мозг вызывает неприятные ощущения - синдром похмелья; далее ацетальдегид постепенно окисляется митохондриальной ацетальдегиддегидрогеназой и образующийся ацетат может служить для синтеза жирных кислот и т.п. Ощущения, вызываемые ацетальдегидом, явились отправной точкой для создания ряда противоалкогольных средств, подавляющих ацетальдегиддегидрогеназу, повышающих тем самым уровень, ацетальдегида и ускоряющих, в результате, развитие тяжелого состояния после приема даже небольших доз алкоголя. В практику вошел, в частности, такой ингибитор этого фермента, как тетурам, систематическое введение которого приносит определенную пользу в лечении алкоголизма. Особенно перспективными оказались воздействия, позволяющие на длительное время изменять активность двух указанных главных ферментов метаболизма этанола. В экспериментах на крысах-алкоголиках эффективной оказалась индукция аутоантител, связывающих эти ферменты.
а взаимодействуя с серотонином, - метил-тетрагидро- -карбо-лин:
Эти соединения имеют некоторое структурное сходство с морфином.
Адетальдегид способен также тормозить один из этапов катаболизма дофамина - его окислительное дезаминирование, - так что накапливается промежуточный продукт - 3,4 - д иокси-фенилацетатальдегид. Последний, взаимодействуя опять-таки с дофамином, образует тетрагидропапаверолин, способный, в свою очередь, превращаться в соединения, все более приближающиеся по структуре к морфину, в том числе - норморфин:
В последние годы прослежены метаболические пути, ведущие к образованию в организме млекопитающих даже кодеина и морфина, хотя и в очень малых количествах. Более того, сейчас можно считать, что в микроконцентрациях многие соединения этого ряда постоянно представлены в мозге. Однако введение извне этанола и образование из него ацетальдегида резко повышает уровень морфиноподобных соединений.
Установлена способность сальсолинола и других эндогенных аналогов морфина, образующихся с участием ацетальдегида, служить как агонистами, так и блокаторами опиоидных рецепторов в зависимости от концентрации и других условий. Следствия такого взаимодействия могут состоять, во-первых, в подмене эндогенных факторов вознаграждения и, во-вторых, если концентрация сальсолинола в организме алкоголика постоянно повышена, то блокада рецепторов в отношении собственных эндогенных, наиболее адекватных, факторов вознаграждения может вызвать постоянное чувство неудовлетворенности и побуждать к поиску наркотических средств.
Пока трудно отдать предпочтение одной из этих возможностей, но образование сальсолинола и подобных ему непептидных морфиноподобных соединений при алкоголизме указывает на вероятную связь опиоидной системы с механизмом алкоголизма. К этой же мысли приводит тот факт, что классический блокатор опиатных рецепторов - налоксон оказался полезен также при лечении алкоголизма.
Наконец, показательно, что у большинства алкоголиков возрастает уровень антител к морфиноподобным соединениям. Понятна поэтому настойчивость, с которой современные исследователи после открытия опиоидных пептидов ищут корреляции между уровнем последних, а также состоянием опиоидных рецепторов, с одной стороны, и глубиной и фазой алкоголизма, с другой стороны. Найденные сейчас корреляции, подтверждающие более или менее значительное участие системы опиоидов в механизмах алкоголизма, свидетельствуют о довольно сложных отношениях, подчас противоречивых. Так, например, показано меньшее содержание метэнкефалина в мозге предрасположенных к алкоголю животных и меньшая концентрация в гипоталамусе р-эндорфина у животных со сформированным алкоголизмом и наследственно предрасположенных к алкоголизму.
Установлено также, хотя и не на всех экспериментальных моделях, что введение таким животным этанола повышает уровень метэнкефалина и р-эндорфина.
Можно пытаться толковать эти данные так, что сниженные уровни эндогенных опиоидов в мозге обусловливают влечение к этанолу как к фактору, ведущему к образованию в мозге опиоидов, т.е. к нормализации гуморальных систем вознаграждения. С этим согласуются феномены снятия абстиненции и некоторого снижения влечения к алкоголю при введении извне опиоидных нейропептидов, а также некоторых ингибиторов протеолитического распада опиоидных пептидов в организме. Однако ряд экспериментальных данных трудно согласовать с таким толкованием. Так, в отличие от метэнкефалина содержание лейэнкефалина в мозге предрасположенных к алкоголизму животных повышено.
Противоречивы данные об изменениях уровня р-эндорфина в плазме и цереброспинальной жидкости при введении этанола. Источником недоразумений является также то, что при алкоголизме возможны, и в ряде работ зарегистрированы, не только изменения уровня эндогенных опиоидов, но и состояния их рецепторов в силу упоминавшейся мембранотропности этанола. Есть данные о снижении сродства энкефалинов к их рецепторам под действием этанола. Следовательно, для строгого учета роли эндогенных опиоидов при алкоголизме необходимо совместное рассмотрение данных об уровне опиоидов и о состоянии их рецепторов. Таких данных пока недостаточно.
Богатый, хотя опять-таки неоднозначный, экспериментальный материал собран о роли катехоламинов и серотонина в развитии алкоголизма. Здесь прослеживается четкая зависимость действия этанола от этапа, фазы развития алкоголизма.
Однократный прием этанола вызывает вначале усиленный выброс катехоламинов - дофамина и норадреналина, обусловленный, вероятно, мембранотропным действием алкоголя на пресинаптические рецепторы. Секретируемые катехоламины входят в число факторов внутреннего вознаграждения, вызывающих эйфорическое состояние. После выброса катехоламинов срабатывает система обратной регуляции, которая не просто нормализует состояние системы, а создает временный дефицит катехоламинов в синаптической щели и жидкостях организма. Возможно, это служит одним из побуждающих факторов к повторению приема этанола.
При развитии хронического алкоголизма состояние сниженного выхода, усиленной деградации и повышенного обратного захвата катехоламинов как бы закрепляется, создавая, как полагают, постоянный механизм, побуждающий к частому приему этанола для временной коррекции этих нарушений. Полное прекращение приема алкоголя на стадии развитого хронического алкоголизма ведет к экстренной мобилизации всех существующих механизмов синтеза, выброса и сохранения катехоламинов в синаптической щели. Разрегулированная на предыдущих стадиях система срабатывает так, что происходит не нормализация уровня катехоламинов, а чрезмерное возрастание их концентрации, в частности дофамина. Они участвуют в развитии абстиненции.
Сходной является, в общем, и динамика изменений выхода и превращений серотонина. Извращения его выброса могут быть связаны с эйфорией и галлюцинациями. Вместе с тем роль серотонинергической системы представляется пока неоднозначной - Так, с одной стороны, ряд стимуляторов синтеза и выхода серотонина, блокаторы обратного захвата и многие агонисты подавляют влечение к этанолу. В то же время селективные антагонисты 5НТ 3 -рецепторов и такие ингибиторы синтеза серотонина, как р-хлорфенилаланин, также подавляют потребление этанола. Активная иммунизация животных против серотонина ведет к снижению его уровня в плазме крови и в мозге и к подавлению влечения к алкоголю экспериментальных животных. По-видимому, участие серотонинергической системы в механизмах влечения к алкоголю очень тесно связано с типом рецепторов серотонина, а также локализацией как рецепторов, так и мест синтеза этого медиатора.
В механизмы наркоманий и алкоголизма вовлечена также главная тормозная система мозга - ГАМК-ергическая. Подавление этой системы позволяет понять устойчивость патологических влечении.
Характерно, что действие на влечение к алкоголю ряда нейропептидов более или менее коррелирует с их участием в развитии или подавлении стрессовых состояний. Вообще известно, что стресс сам по себе стимулирует влечение к алкоголю. Пептид дельтасна, оказавшийся сильным противострессовым агентом, достоверно снижает потребление этанола экспериментальными животными при систематическом его введении. У предрасположенных к алкоголю белых крыс его содержание в плазме крови и стриатуме снижено.
В целом представленные результаты исследований в области нейрохимических механизмов алкоголизма не образуют пока единой стройной картины, но свидетельствуют об относительной близости времени, когда она сформируется.
3. СТРАХ, ФОБИИ, р -КАРБОЛИНЫ, ЭНДОЗЕПИНЫ И ХОЛЕЦИСТОКИНИН _4
Сравнительно давно фармакологи создали новый класс транквилизаторов -бензодиазепины, вошедшие сейчас в широкую медицинскую практику. Затем были выявлены рецепторы этих соединений в головном мозге. Поскольку не были известны внутренние лиганды этих рецепторов, их обозначили как рецепторы диазепама. Далее, оказалось, что эти рецепторы являются частью рецепторов гамма-аминомасляной кислоты или самостоятельным рецептором, прочно связанным с рецептором ГАМК. Наконец, удалось выделить часть эндогенных лигандов этих рецепторов: во-первых, большой пептид - эвдозепин, состоящий примерно из сотни аминокислотных остатков, его активные фрагменты - малые 18- и 6_членные пептиды и, во-вторых, непептидные соединения - производные так называемых -карболинов. Примером последних является метилтетрагидро -карболин . Активные фрагменты эндозепинов имеют структуру: QATVGDVNTDRPGLLDLK и GLLDLK.
Эти соединения оказывают действие на поведение животных, обратное действию ГАМК и ее аналогов. Они вызывают беспокойство, проявления страха и в опытах на грызунах проконфликтное поведение. В США документировано острое беспокойство, паническое состояние людей, которым вводили одно из производных -карболина.
Что касается транквилизаторов - бензодиазепинов, с которых начался этот цикл исследований, то они оказались блока - торами рецепторов эндозепинов, подавляющими их взаимодействие с эндогенными факторами страха, беспокойства и проконфликтного поведения.
В последние годы внимание нейрохимиков и психиатров привлек еще один пептид, вызывающий беспокойство, страх и паническое поведение как у людей, так и у животных, - наименьший из обнаруживаемых в мозге С-конпевых фрагментов холецистокинина - ХЦК_4. Его действие на поведение опосредовано стимуляцией некоторых отделов дофаминергической системы через специальные рецепторы ХЦК В. Уже синтезированы антагонисты ХЦК_4, с помощью которых удается снизить уровень тревожности и панических реакций как в опытах на животных, так и в первых клинических исследованиях.
Для понимания биохимических механизмов ряда расстройств психики значение этих открытий весьма велико. Многие психические расстройства сопровождаются навязчивыми страхами, фобиями, крайне беспокойным и конфликтным поведением. Они характерны, в частности, для поздних стадий алкоголизма, некоторых проявлений шизофрении и др.
4. ДОФАМИН И ПАРКИНСОНИЗМ
Раскрытие биохимических процессов, лежащих в основе болезни Паркинсона - глубокого нарушения стереотипной двигательной активности, ее координации и инициации, - стало одним из первых ярких достижений патологической нейрохимии.
Синдром болезни удалось воспроизвести в экспериментах на животных, вводя им 6 _о ксидофамин . Этот аналог дофамина проникает в везикулы нервных окончаний, предназначенные для накопления и выброса катехоламинов, конкурирует с последними за включение в везикулы и, в конечном счете, подавляет катехоламинергическую трансмиссию. Этот процесс иногда называют химической десимпатизацией, имея в виду особую роль катехоламинов в симпатической нервной системе. Однако это название неточно, ибо катехоламины широко распространены и функционируют во многих других отделах нервной системы.
Дофаминергические нейроны стриатума, части хвостатого ядра и особенно черной субстанции, являющиеся основными центральными организаторами стереотипной двигательной активности, оказались высокочувствительными к такому действию 6_оксидофамина. В результате впервые удалось, вводя вещество определенной биохимической направленности действия, вызвать такое специфическое заболевание, как паркинсонизм.
В последние годы появилась возможность еще более точного определения нейронов, повреждение которых достаточно для возникновения паркинсонизма. Синтетический нейротоксин - метилфенилтетрапэдропиридин избирательно связывается с меланинсодержащими нейронами черной субстанции, вызывая их депигментацию и паркинсонический синдром. Отмечено также существенное снижение содержания в черном веществе метэнкефалнна и холецистокинина и обнаружен, наконец, дефицит одного из глиальных белков, выполняющих тропические функции по отношению к нейронам, синтезирующим глутамин.
Участие дофаминергических систем в паркинсоническом синдроме предполагает возможность облегчения синдрома введением в мозг дофамина. Поскольку дофамин не проходит гематоэнцефалический барьер, воспользовались для введения больным его ближайшим предшественником - диоксифенилаланином. Он существенно облегчает состояние паркинсоников. Следует, однако, подчеркнуть, что длительное введение больным больших доз диоксифенилаланина, значительно усиливающего синтез дофамина во всех отделах мозга, может вести к появлению симптомов, сходных с другим психическим заболеванием - шизофренией, одним из физиологических и биохимических проявлений которого является именно гиперактивность дофаминергической системы.
Новейший путь лечения паркинсонизма состоит в пересадках клеток или участков ткани здорового мозга из мезэнцефалона человеческих плодов, способных продуцировать дофамин, в определенные участки мозга больного. Заметим, что особенности иммунологического статуса мозга значительно облегчают такие пересадки. Результаты первых серий таких пересадок обнадеживают. Сейчас накапливается опыт длительного наблюдения следствий этих операций и разрабатываются культуры клеток мезэнцефалона с тем, чтобы отказаться от использования материалов, получаемых при абортах.
5. ШИЗОФРЕНИЯ, КАТЕХОЛАМИНЫ И ВНУТРЕННИЕ НЕЙРОЛЕПТИКИ
Практически полный отказ современной психиатрии от палат для буйных психических больных объясняется двумя причинами, различными внешне, но сходными по сути. Первая состоит в хирургическом вмешательстве - перерезании катехол-аминергических путей, идущих к лобной коре от таламуса, ретикулярной формации, черной субстранции и некоторых других отделов мозга. Довольно эффективное для снятия агрессивных проявлений шизофрении, это средство, тем не менее, связано с определенной деградацией умственных способностей и в настоящее время уступило место фармакологическим воздействиям. Последние состоят в подавлении рецепции и/или секреции катехоламинов, особенно дофамина, такими соединениями, как галоперидол, трициклические нейролептики и др. Строго говоря, эти агенты не столько лечат больных от шизофрении, сколько подавляют ее проявление: агрессивное поведение, галлюцинации, стереотипную двигательную активность и т.п.
Эффективность указанных средств приближает к пониманию механизмов болезни, включающих глубокое извращение и патологическое усиление в определенных отдела, мозга шизофреников катехоламинергической, особенно дофал. инергической, трансмиссии. Установлено 4-5_кратное повышение плотности рецепторов дофамина D 4 . Показательно, что одно из лучших антипсихотических лекарств - клозапин - обладает наибольшим сродством именно к рецепторам D 4 . Выявлен также значительно повышенный уровень дофамина в височной доле головного мозга, особенно в левой миндалине. Отмечен и ряд морфологических изменений в тех же отделах - увеличение объема боковых желудочков, утончение парагиппокампальной коры и др.
Противоречивыми, в отличие от закономерных изменений содержания дофамина, являются сведения о содержании норадреналина в различных отделах мозга шизофреника. Относительно воспроизводимы лишь данные о повышении уровня норадреналина в цереброспинальной жидкости.
Особого внимания заслуживают сообщения о воспроизведении отдельных проявлений шизофрении при воздействии агентов, так или иначе вмешивающихся в состояние катехоламинергической и серотонинергической систем. Так, аналог дофамина - растительный алкалоид мецкалин вызывает галлюцинации, в том числе цветные, имеющие сходные элементы с шизофреническими.
Животные, получившие высокие дозы фенамина, после периода возбуждения проявляют монотонную, стереотипную двигательную активность, напоминающую таковую у шизофреников.
Галлюцинаторные явления, наблюдаемые при введении некоторых аналогов серотонина, например диэтиламида лизергиновой кислоты , также заслуживают внимания с точки зрения возможной роли при шизофрении не только извращений катехоламинергической, но и серотонинергической трансмиссии.
Картина патологического, несбалансированного усиления при шизофрении катехоламинергических и, возможно, серотонинергических систем в определенных участках мозга хорошо согласуется и с данными об изменении набора и активности моноаминооксидаз, расщепляющих соответствующие нейромедиаторы после выхода из нервного окончания. Из четырех форм МАО, обнаруженных в мозге человека, - I, II, IIи III - у шизофреников отсутствует III и существенно модифицирована II форма.
Наконец, в механизмы шизофрении вовлечена еще одна нейромедиаторная система - глутаматергическая. Отмечено ее значительное ослабление во фронтальной коре. Напомним, что с этой системой особенно тесно связан ряд высших функций мозга. Такой антагонист глутамата, как пенциклидин, имитирует некоторые симптомы шизофрении на животных. Сейчас изучается возможность применения агонистов глутаматергических рецепторов для терапии шизофрении.
Меньше данных накоплено пока в отношении изменений пептидной регуляции при шизофрении.
В нервных окончаниях, как правило, встречаются те или иные ассоциации классических яюйромедиаторов с нейропептидами. В частности, дофаминергические окончания особенно часто содержат холецистокинин _8, вазоактивный интестинальный пептид и, реже, соматостатин и вещество Р. В норад-рененргических окончаниях частыми спутниками норадреналина являются нейропептиды Y и энкефалины. Известно также, что холецистокинин_8 и, с меньшей определенностью, соматостатин тормозят дофаминергическую передачу. Аналогичные данные имеются в отношении действия энкефалинов на норад-ренергическую трансмиссию. Активатором же норадренергаческой
передачи является нейропептид Y, а дофаминергической, опять-таки с меньшей определенностью, - вазоактивный интестинальный пептид. Поэтому существенное значение имеют данные о снижении содержания холецистокинина_8 в гиппокампе и миндалине мозга шизофреников. Снижено также содержание в гиппокампе соматостатина, а в миндалине - метэнкефалина и вещества Р, однако концентрация вазоактивного интестинального пептида в миндалине повышена.
Со всей этой группой данных ассоциируются сведения о нейролептическом действии холецистокинина_8 и родственного ему пептида - церулеина, а также некоторых эндорфинов. Оно проявляется в тестах, разработанных в свое время для отбора лекарств-нейролептиков: по способности вызывать каталепсию, по действию на агрессивное поведение, по подавлению эффектов фенамина и др. Снижение ими дофаминергической трансмиссии показано и в экспериментах на клеточном уровне. Все это послужило основанием для наименования холецистокинина_8, а также дезтирозил- и дезэнкефалин-эндорфинов эндогенными нейролептиками. К сожалению, попытки использовать их как лечебные средства для купирования шизофренического синдрома у людей пока не дали ощутимых результатов. Можно говорить пока лишь о положительных тенденциях, в частности о перспективности совместного применения холецистокинина или церулеина с обычными нейролептиками.
Более перспективными представляются исследования, основанные на данных о способности короткого концевого тетрапептида холецистокинина - ХЦК_4 - вызывать состояния тревоги, страха и паники. Действие ХЦК_4 опять-таки связано с активацией части дофаминергической системы. Поэтому на синтезе и испытаниях антагонистов ХЦК_4 основаны надежды на создание нового класса лекарственных веществ для лечения некоторых форм шизофрении.
6. КАТЕХОЛАМИНЫ, НЕЙРОПЕПТИДЫ И ДЕПРЕССИВНЫЕ СОСТОЯНИЯ
Подавление катехоламинергической и серотонинергической систем сопряжено с депрессивным состоянием. Однако картина подавления катехоламинергической трансмиссии при депрессиях не является простым обращением того, что наблюдается, например, при агрессивных формах шизофрении, а механизмы депрессий не сводятся только к нарушениям классических медиаторных систем.
Накопилось немало данных о существенном снижении норадренергической трансмиссии при депрессиях. В меньшей степени снижается активность серотонинергической системы. Еще меньше сдвигов в дофаминергической трансмиссии. Отсутствуют существенные изменения в других классических нейромедиаторных системах. В согласии с этими данными находится эффективность снятия депрессии большой группой лекарственных средств - имипрамином, амитршггилином, инказаном и многими другими, действие которых состоит либо в торможении обратного захвата норадреналина и серотонина, либо в ингибировании их расщепления моноаминооксидазами. Заметим лишь, что таков механизм действия антидепрессантов только на начальных этапах применения. При длительном же их введении включаются сложные изменения в рецепции медиаторов. При введении людям веществ, снижающих уровень катехоламинов, может возникнуть депрессия. Таким веществом оказался резерпин, применяемый при лечении гипертонии.
Таким образом, значимость снижения активности катехоламинергической системы при развитии депрессий очевидна; однако это снижение не является простой противоположностью тем изменениям, которые описаны для шизофрении. При шизофрении - преобладающее усиление дофаминергической трансмиссии, при депрессии - преимущественное подавление норадренергической и серотонинергической. Полагают также, что в характерном для депрессий подавлении психической и двигательной активности участвует также дисбаланс между возбуждающей глутаматергической и тормозной ГАМК-ергической системами, отмечаемый как при депрессиях, так и при шизофрении. Наконец, в мозге больных депрессиями обнаружены значительные, изменения плотности опиатных рецепторов. Это указывает на возможные отклонения в системе «внутреннего вознаграждения».
7. СУДОРОЖНЫЕ СОСТОЯНИЯ, ЭПИЛЕПСИЯ, ГЛУТАМАТНЫЕ РЕЦЕПТОРЫ И ИХ АНТАГОНИСТЫ
Биохимические механизмы судорожных состояний и особенно такой болезни, как эпилепсия, принципиально отличаются от описанных выше для шизофрении, депрессий, наркоманий и алкоголизма. Важная роль в индукции синдрома эпилепсии принадлежит глутаминергической системе. Очевидно также значение ГАМК-ергической и эндозепамергической систем.
С глутаминергической трансмиссией тесно связана не только возможность индукции судорожных состояний, но и ряд высших функций ЦНС, таких, например, как память. В этом одна из причин того, что эпилепсия не сводится к судорожному синдрому и сопряжена с рядом сложных изменений психики.
Рецепторы глутаминовой кислоты - образования сложные и неоднородные. Характерно, что раскрытие их структуры и разнообразия было ускорено обнаружением веществ, которые иногда называют возбуждающими нейротоксинами. К ним относится каиновая кислота, квисквалевая кислота и ряд других соединений, многие из которых имеют общие с глутаматом элементы структуры. Сам по себе глутамат при интрацеребральном введении в определенные зоны мозга может вызывать приступы судорог. Однако каи-нат и квисквалат оказались особенно мощными индукторами судорог и, более того, агентами, способными специфически разрушать нейроны, несущие глутаматные рецепторы.
На глутаматных рецепторах выявлены участки связывания барбитуратов - агентов, тормозящих их функцию и обладающих соответственно противосудорожной активностью. Один из самых мощных и специфичных блокаторов NMDA_глутаматных рецепторов - 2 _а мино _7_ф осфоногептановая кислота - предотвращает припадки эпилепсии у экспериментальных животных. Все это заставляет считать изменения глутаминергической трансмиссии одними из узловых в патогенезе эпилептиморфных судорожных состояний.
Поддержкой этой же гипотезы служит обнаружение в плазме крови эпилептиков значительно повышенных уровней антител к белкам глутаматного рецептора. Это используется для диагностики скрытых форм эпилепсии и оценки тяжести заболевания. Подобное явление отражает, по-видимому, снижение функций гематоэнцефалического барьера при развитии эпилепсии, сопровождающееся выходом в периферический кровоток определенных количеств белков рецептора и их фрагментов, и, далее, образованием антител к ним. Возможно, сам патогенез эпилепсии подобен патогенезу аутоиммунных болезней мозга, при которых аутоантитела к белкам мозга служат основным повреждающим фактором. В том, что последняя гипотеза правомочна, убеждают, во-первых, данные о повышенном уровне в плазме крови эпилептиков не только антител к белкам глутаматного рецептора, но и к другим белкам и липидам синаптических мембран мозга, например к белку S_100, а также результаты экспериментов с введением в мозг антител, полученных к различным белковыми и липидным фракциям мозга. Эпилептиформные процессы возникали, в частности, при инъекции антител к некоторым фракциям ганглиозидов.
Другая система, связь которой с судорожными состояниями и эпилепсией весьма вероятна, - ГАМК-ергическая.
Тормозные функции ГАМК-ергической системы носят более универсальный, менее специфический характер, чем функции возбуждающих нейромедиаторных систем. Это отражает, в частности, тот факт, что доля ГАМК-ергических терминалей в мозге является наибольшей. Снижение судорожной готовности и облегчение судорожных состояний установлено при центральном введении ГАМК, а также при периферическом введении его аналогов, способных проходить через гематоэнцефалический барьер. Таков же эффект соединений, тормозящих распад, стимулирующих синтез и обратный захват ГАМК: вальпроата натрия , а также прогабида, у-ацетилен-ТАМК и др.
На рецепторах ГАМК обнаружен барбитурат-связывающий участок. В этом случае в отличие от аналогичного участка на глутаматных рецепторах барбитураты усиливают эффект основного нейромедиатора.
Таким образом, барбитураты проявляют противосудорожное действие, изменяя состояние двух категорий рецепторов: глутаматных, подавляя возбуждающее действие ихлигандов, и ГАМКд-рецепторов, стимулируя их тормозное действие.
Еще более подтверждает представление о роли ГАМК-ергической системы в предотвращении и купировании судорожных состояний тот факт, что подавление синтеза ГАМК, напротив, провоцирует судороги. Ярким примером являются судороги у людей при авитаминозе В 6 . Витамин В 6 служит предшественникам пиридоксаль_5_фосфата, который, в свою очередь, является кофактором глутаматдекарбоксилазы, катализирующей последнюю стадию синтеза ГАМК. Провоцируют судороги и такие ингибиторы этого фермента, как аллилглицин, гидразиды и др. Судорожные припадки вызывает и один из ядов грибов - п икротоксинин, тоже связывающийся с ГАМК-рецепторами и подавляющий их активность. С ГАМК-рецепторами сопряжены, участки связывания эндозепинов - пептидов, вызывающих возбуждение, страх и проконфликтное поведение. Они снижают активность ГАМК-рецепторов. Понятно поэтому, что блокаторы рецепторов эндозепинов - бензодиазепиновые транквилизаторы - оказались агентами, облегчающими эпилептические процессы.
В этой связи, хотя излагаемый материал касается природы эпилепсии и эпилептических судорог, следует отметить и данные о патогенезе судорог, вызываемых такими ядами, как стрихнин и столбнячный токсин. Стрихнин блокирует рецепторы глицина, второго по значимости после ГАМК тормозного медиатора центральной нервной системы, функционирующего преимущественно в спинном мозге. Действие же столбнячного токсина направлено преимущественно на блокаду выхода ГАМК из нервных окончаний в головном мозге, что ведет к блокаде тормозных влияний на мотонейроны. Эти данные опять-таки подчеркивают роль нарушений систем тормозных нейромедиаторов.
Была бы, однако, ошибочной попытка свести патогенез эпилепсии и ряда других судорожных состояний только к изменениям глутаминергической системы или ее баланса с ГАМК-ергической системой. Яркой иллюстрацией сложности проблемы и недопустимости поспешного формирования гипотез в этой области являются, например, данные о способности опиоидного пептида - метэнкефалина - вызывать судороги при аппликации в некоторых участках мозга - вне зон, где он проявляет аналитическое действие, типичное для опиоидов. Эти данные послужили также основой для предположений, что эпилептический приступ - результат чрезмерно высокого выброса энкефалина из опиоидергических терминалей с распространением его на зоны мозга, где он способен индуцировать судороги. Предэпилептическое состояние - аура - истолковывалось при этом как эйфоригенное действие опиоида. К сожалению, вся известная совокупность данных о механизмах эпилепсии не позволяет принять столь простое толкование. Здесь особенно важно сочетанное исследование биохимических и физиологических механизмов эпилепсии.
Особого внимания заслуживают появляющиеся в последнее время данные о роли пептидергических систем при эпилепсии и о существовании эндогенных пептидных лигандов глутаматных рецепторов.
Патогенез судорожных состояний и эпилепсии в отличие от многих других психических болезней тесно связан с изменениями энергетических процессов в нейронах, причем в тех из них, которые входят в патологические эпилептогенные очаги.
Отмечены изменения метаболической структуры нейронов и их митохондрий: повышение проницаемости мембран для К + и Na + , коррелирующее с повышением чувствительности белков мембран к протеиназам, снижение синтеза АТФ и др. Аномальные локальные изменения концентраций К + и Na + , а также NH 4 + , выделяющегося при эпилептическом приступе в результате усиления реакций дезамидинирования, могут вызывать или облегчать деполяризацию постсинаптических зон, снижать порог возбудимости и провоцировать судорожный приступ.
С поддержанием энергетического статуса мозга тесно связана система аденозиновых рецепторов Aj. Эти рецепторы выполняют ряд тормозных энергосберегающих функций, вызывая седативные и противосудорожные эффекты. Включение их осуществляется аденозином и аденозинмонофосфатом - конечным продуктом утилизации таких макроэргов, как АТФ и АМФ. Показаны изменения уровня этих соединений при эпилепсии и изучается возможность создания на этой основе новых противоэпилептических средств.
Сейчас трудно окончательно решить вопрос о первичности нарушений в медиаторных, рецепторных или энергетических системах в патогенезе эпилепсии. Несомненна, однако, их взаимодействие. На это указывают глубокие локальные изменения энергетических систем при инъекции каиновой кислоты именно в патологически измененных участках «эпилептического» мозга.
8. ХОЛИНЕРГИЧЕСКИЕ СИСТЕМЫ МОЗГА, р-АМИЛОИД, НЕЙРОПЕПТИДЫ И СЕНИЛЬНЫЕ ДЕМЕНЦИИ
Разновидность старческого слабоумия, так называемая сенильная деменция Альтцгеймеровского типа, сопровождается прежде всего дегенерацией большого числа подкорковых нейронов, холинергические терминали которых широко распространены в мозге. Далее возникают дегенеративные процессы в м-холинергических системах коры и гиппокампа, сопровождаемые снижением уровня ключевого энзима синтеза ацетилхолина - холинацетилтрансферазы и характерными морфологическими изменениями: дефицитом крупных пирамидных нейронов, появлением скоплений белка - участвует в межнейронных контактах и 2) частично расщепляясь специфическими протеазами, образует большой N_концевой фрагмент, который выходит во внеклеточные среды мозга и принимает участие в процессах консолидации памяти. При болезни Альтцгеймера искажается протеолиз белка-предшественника и из средней его части вышепляется небольшой фрагмент из 41 аминокислотного остатка - собственно р-амилоид, откладывающийся на поверхности нейронов. Перестает формироваться и упомянутый выше большой фрагмент, стимулирующий консолидацию памяти. В результате возникает глубокое нарушение способности больных к запоминанию.
Дегенерация больших групп нейронов, входящих в м-холинергическую систему, сопровождается также глубокими изменениями ряда нейропептидных систем. К сожалению, пока нельзя сказать, какие из них первичны, а какие вторичны, но очевидно очень значительное снижение уровней кортиколиберина в затылочной коре и в хвостатом ядре, а также соматостатина в височной и лобной коре. В то же время возрастает уровень нейропептида Y в так называемой безымянной субстанции.
В отличие от ряда других психических болезней пока мало информации о лекарственных средствах, облегчающих сенильную деменцию, и поэтому мало соответствующих дополнительных косвенных указаний на биохимические механизмы болезни. Можно лишь отметить имеющиеся неравноценные данные о кратковременно облегчающих болезнь средствах, усиливающих холинергическую трансмиссию, что, во всяком случае, не противоречит изложенным выше представлениям.
В порядке иллюстрации того, как вредна поспешность в формировании научных гипотез, следует упомянуть предположение о том, что причиной болезни Альтцгеймера является токсическое действие алюминия. Авторы гипотезы исходили из сообщений о высоком содержании алюминия в нейронах гиппокампа и некоторых других отделах мозга у людей, страдавших болезнью Альтцгеймера. Этот факт был сопоставлен с широким употреблением алюминиевой посуды и содержащих алюминий пищевых добавок в современную историческую эпоху. Однако, по данным последних работ в этой области, изменения содержания алюминия если и существуют, то не столь велики, как казалось ранее, причем есть основания полагать, что проникновение в мозг и связывание нейронами алюминия - это вторичный процесс, являющийся следствием нарушения защитной функции гематоэнцефалического барьера-
9 . КАТИОННЫЙ БЕЛОК МИЕЛИНА, НАРУШЕНИЯ ИММУНОЛОГИЧЕСКОЙ АВТОНОМИИ МОЗГА, АЛЛЕРГИЧЕСКИЙ ЭНЦЕФАЛОМИЕЛИТ И РАССЕЯННЫЙ СКЛЕРОЗ
Значительным достижением нейрохимии и нейроиммунологии является раскрытие ведущих механизмов ряда заболеваний, в основе которых лежит процесс повреждения и частичного рассасывания миелиновой оболочки аксонов, в том числе головного мозга. Этот процесс характерен для таких заболеваний, как аллергический энцефалит и рассеянный склероз, и сопровождается глубокими нарушениями функций ЦНС, в частности анорексией, атаксией и параличами; завершается он в значительной доле случаев летально.
Ключевым экспериментом послужило открытие возможности вызывать аллергический энцефалит у разнообразных экспериментальных животных, если вводить им периферически собственный катионный белок миелина с иммуностимулятором - адъювантом Фрейнда.
КБМ составляет около трети белков миелина. и представляет собой крупный пептид, включающий 160-190 а.о., среди которых особенно велика доля остатков основных аминокислот - лизина, аргинина и гистидина - около 25% суммарно. Даже однократное введение этого белка в дозах менее 0,1 мг/кг с иммуностимуляторами вело через 2-4 недели к развитию у большей части животных аллергического энцефалита. Были выявлены, далее, несколько участков КБМ, ответственных за этот эффект. Минимальными по размеру оказались FSWGAEGQR и GSLPQKAQRPQDEN, энцефалитогенность которых близка или даже превышает исходный КБМ.
В конечном счете сложилось представление о том, что катионный белок миелина или его фрагменты, вводимые животным системно или поступающие из мозга в общий кровоток в результате какого-то повреждения гематоэнцефалического барьера, могут восприниматься внемозговой иммунной системой как чужеродные при условии одновременной иммуностимуляции. Возможность восприятия специфических белков центральной нервной системы как чужеродных является следствием частичной иммунологической автономии мозга, возникающей еще в эмбриогенезе. Поэтому поступление катионного белка миелина или его фрагментов на фоне иммуностимуляции ведет к развитию против них иммунного ответа. В частности, происходит специфическая активация Т-лимфоцитов; процесс завершается привлечением и активацией макрофагов, которые и повреждают миелин.
Поддержкой гипотезы об иммунных механизмах аллергического энцефалита служат данные о возможности его профилактики введением больших доз тех же фрагментов катионного белка миелина, но без иммуностимуляторов. Хотя удовлетворительное объяснение механизма возникающей толерантности - задача трудная, очевидна специфичность явления, ибо ряд пептидов и белков, близких по содержанию основных аминокислот не дают профилактического эффекта.
Раскрытие иммунохимических основ аллергического энцефалита и близких к нему болезней - рассеянного склероза и энцефалита, возникающего иногда после вакцинации против вируса бешенства, - заставляет исследователей с возрастающим вниманием относиться к данным об иммунохимических процессах при других психонейрологических болезнях. Выше уже упоминались предположения о возможной роли этих процессов в патогенезе эпилепсии. Разнообразные спектры антител к мозгоспецифическим белкам и ряд изменений иммуно-компетентных клеток регистрируются при шизофрении, депрессиях и др. Очевидной представляется важность накопления и обобщения данных, находящихся на стыке нейрохимии и нейроиммунологии.
В ЫВОДЫ
1. Один из механизмов наркоманий и алкоголизма состоит в имитации наркотиком, этанолом или их метаболитами функций эндогенных соединений, являющихся в норме факторами «внутреннего подкрепления», «вознаграждения» и т.п. Воздействие на их образование и рецепцию - перспективный путь преодоления состояний абстиненции и зависимости.
2. Главные ферменты метаболизма этанола - алкогольдегидрогеназа и ацетальдегиддегидрогеназа. Модуляция их активности оказывает влияние на формирование и поддержание зависимости от алкоголя.
3. Основные нейрохимические механизмы болезни Паркинсона состоят в подавлении синтеза дофамина клетками черной субстанции мозга.
4. Важными элементами нейрохимических механизмов шизофрении являются: усиление синтеза дофамина в височных и некоторых других областях мозга, усиление и извращение катехоламинергической импульсации в направлении от ряда структур среднего мозга к лобной коре, извращение систем удаления и окислительного расщепления моноаминов, изменение пептидергической трансмиссии. Факторы, подавляющие первые два из указанных процессов, временно приостанавливают проявления шизофрении.
5. В основе депрессивных состояний лежит понижение катехоламинергической трансмиссии. Определенную роль играет и извращение серотонинергической системы. Факторы, подавляющие обратный захват и распад катехоламинов, препятствуют проявлению депрессий.
6. Важным элементом механизмов тревожности, патологического страха и проконфликтного поведения может быть активация систем, генерирующих и рецептирующих эндозепины и производные бета-карболина.
7. Характерной чертой патогенеза эпилепсии и ряда других судорожных состояний является повышение и извращение эффективности глутаминергической трансмиссии и нарушения в ГАМК-ергической системе. Факторы, подавляющие первую систему и особенно активирующие вторую, способны купировать судорожные синдромы и некоторые другие проявления эпилепсии.
8. Основой проявлений ряда старческих психозов является нарушение холинергической трансмиссии. Они сопряжены также с извращением синтеза и патологическим накоплением одного из белков ЦНС - бета-амилоида.
9. Многие болезни мозга связаны с иммунохимическими процессами, в том числе с нарушением иммунной автономии мозга. Демиелинизирующие болезни - аллергический энцефалит, рассеянный склероз и др. - могут быть вызваны иммунизацией к катионному белку миелина. При эпилепсии, шизофрении и некоторых других психических болезнях в крови обнаруживаются антитела к отдельным нейроспецифическим белкам.
Подобные документы
Сущность и последствия рассеянного склероза, происхождение его названия. Возникновение заболевания в результате нарушения иммунной системы; история его изучения и факторы, способствующие появлению. Наиболее распространенные симптомы рассеянного склероза.
презентация , добавлен 03.12.2012
Описание рассеянного склероза. Особенности и причины возникновения, появление бляшек в спинном и головном мозге. Наследственная предрасположенность. Клинические признаки. Атактический синдром. Нарушение чувствительности. Типы течения рассеянного склероза.
презентация , добавлен 21.03.2017
Класс препаратов, выделенных из макового сока для анальгезии (опиоиды). Эндогенные опиоиды, применение в периоперационном периоде, для регионарной анестезии. Побочный эффект спинального применения опиоидов. Сердечно-сосудистые эффекты опиоидных агонистов.
контрольная работа , добавлен 04.08.2009
Представление схемы нервных волокон головного и спинного мозга. Характеристика ассоциативных, комиссуральных (спаечных) и проекционных типов проводящих путей. Классификация нервных волокон по различных признакам. Черепные и спинномозговые нервы.
презентация , добавлен 27.08.2013
Общая характеристика мозговых механизмов высших психических функций, особенности системного представления о их локализации. Основные методологические положения антилокализационистов. Синдромный анализ нарушения высших психических функций, его факторы.
контрольная работа , добавлен 26.11.2010
реферат , добавлен 21.07.2013
Особая значимость патогенетической терапии в клинике нервных болезней. Типовые патологические процессы в нервной системе. Нарушение нервной трофики. Генераторы патологически усиленного возбуждения. Механизм повреждения нейронов при ишемии мозга.
лекция , добавлен 13.04.2009
Влияние медикаментозных препаратов, используемых для лечения психических болезней, на выбор анестезии. Специфика анестезиологического пособия и осложнения у больных хроническим алкоголизмом, наркоманией, в стадии острого опьянения, при табакокурении.
реферат , добавлен 13.03.2010
Наследственные и приобретенные нарушения обмена веществ. Метаболические энцефалопатии как расстройства различных отделов ЦНС. Нарушения мозгового кровотока, миелина, нервных механизмов управления движениями и нарушение движений при повреждении мозжечка.
реферат , добавлен 13.04.2009
Шизофрения и ее формы. Шизоаффективное расстройство. Онейроидная кататония. Ранняя детская шизофрения, ее симптомы. Факторы риска детской шизофрении. Клинические особенности шизофрении, варианты течения, характер основных расстройств, возможные исходы.
^ 20.биохимические основы качества силы и пути его развития
Под силой мышц обычно понимается способность преодолевать внешнее сопротивление, либо противодействовать ему посредством мышечных напряжений.
Скоростно-силовые качества главным образом зависят от энергообеспечения работающих мышц и от их структурно-морфологических особенностей, в значительной мере предопределенных генетически.
Проявление силы и быстроты характерно для физических нагрузок, выполняемых в зоне максимальной и субмаксимальной мощности. Следовательно, в энергообеспечении скоростно-силовых качеств преимущественно участвуют анаэробные пути ресинтеза АТФ – креатин-фосфатный и гликолитический.
Быстрее всего развертывается ресинтез АТФ за счет креатинфосфатной реакции. Она достигает своего максимума уже через 1–2 с после начала работы. Максимальная мощность этого способа образования АТФ превышает скорость гликолитического и аэробного путей синтеза АТФ в 1,5 и 3 раза соответственно. Именно за счет креатинфосфатного пути ресинтеза АТФ мышечные нагрузки выполняются с самой большой силой и скоростью. В свою очередь, величина максимальной скорости креатинфосфатной реакции зависит от содержания в мышечных клетках креатинфосфата и активности фермента креатинкиназы. Увеличить запасы креатинфосфата и активность креатинкиназы возможно за счет использования физических упражнений, приводящих к быстрому исчерпанию в мышцах креатинфосфата.
Для этой цели используются кратковременные упражнения, выполняемые с предельной мощностью. Хороший эффект дает применение интервального метода тренировки, состоящей из серий таких упражнений. Спортсмену предлагается серия из 4–5 упражнений максимальной мощности продолжительностью 8–10 с. Отдых между упражнениями в каждой серии равен 20–30 с. Продолжительность отдыха между сериями составляет 5–6 мин.
Выполнение скоростных и силовых нагрузок в зоне субмаксимальной мощности обеспечивается энергией в основном за счет гликолитического ресинтеза АТФ. Возможности этого способа получения АТФ обусловлены внутримышечными запасами гликогена, активностью ферментов, участвующих в этом процессе, и резистентностью организма к молочной кислоте, образующейся из гликогена. Поэтому для развития скоростно-силовых способностей, базирующихся на гликолитическом энергообеспечении, применяются тренировки, отвечающие следующим требованиям. Во-первых, тренировка должна приводить к резкому снижению содержания гликогена в мышцах с последующей его суперкомпенсацией. Во-вторых, во время тренировки в мышцах и в крови должна накапливаться молочная кислота для последующего развития резистентности к ней организма.
Промежутки отдыха как между отдельными упражнениями, так и между сериями упражнений явно недостаточны для восстановления запасов гликогена, и вследствие этого в ходе тренировки в мышцах происходит постепенное уменьшение содержания гликогена до очень низких величин, что является обязательным условием возникновения выраженной суперкомпенсации.
Структурно-морфологические особенности мышц, определяющие возможности проявления силы и быстроты, касаются строения как отдельных мышечных волокон, так и мышцы в целом. Скоростно-силовые качества отдельного мышечного волокна зависят от количества сократительных элементов – миофибрилл – и от развития саркоплазматической сети, содержащей ионы кальция. Саркоплазматическая сеть также участвует в проведении нервного импульса внутри мышечной клетки. Содержание миофибрилл и развитие саркоплазматической сети неодинаково в мышечных волокнах разных типов. В зависимости от преобладания тех или иных способов образования АТФ, химического состава и микроскопического строения выделяют три основных типа мышечных волокон: тонические, фазические и переходные. Эти типы волокон также различаются по своей возбудимости, времени, скорости и силе сокращения, продолжительности функционирования.
Тонические волокна содержат относительно большое количество митохондрий, в них много миоглобина, но мало сократительных элементов – миофибрилл. Основной механизм ресинтеза АТФ в таких мышечных волокнах – аэробный. Поэтому они сокращаются медленно, развивают небольшую мощность, но зато могут сокращаться длительное время.
Фазические волокна имеют много миофибрилл, хорошо развитую саркоплазматическую сеть, к ним подходит много нервных окончаний. В них хорошо развиты коллагеновые волокна, что способствует их быстрому расслаблению. В их саркоплазме значительны концентрации креатинфосфата и гликогена, высока активность креатинкиназы и ферментов гликолиза. Относительное количество митохондрий в белых волокнах значительно меньше, содержание миоглобина в них низкое, поэтому они имеют бледную окраску. Обеспечение энергией белых мышечных волокон осуществляется за счет креатинфосфатной реакции и гликолиза. Сочетание анаэробных путей ресинтеза АТФ с большим количеством миофибрилл позволяет волокнам данного типа развивать высокую скорость и силу сокращения. Однако вследствие быстрого исчерпания запасов креатинфосфата и гликогена время работы этих волокон ограничено.
Переходные мышечные волокна по своему строению и свойствам занимают промежуточное положение между тоническими и фазическими.
Даже из такого краткого перечисления различий между типами мышечных волокон следует, что для проявления силы и быстроты более предпочтительны белые волокна и близкие к ним по строению переходные волокна. Поэтому более выраженными скоростно-силовыми качествами, при прочих равных условиях, обладают те мышцы, в которых соотношение между мышечными волокнами смещено в сторону белых.
Соотношение между волокнами разных типов в скелетных мышцах неодинаковое. Так, мышцы предплечья, двуглавая мышца плеча, мышцы головы и другие содержат преимущественно физические волокна. Мышцы туловища, прямая мышца живота, прямая мышца бедра в основном содержат тонические волокна. Отсюда легко понять, почему указанные группы мышц существенно различаются по таким свойствам, как возбудимость, быстрота, сила, выносливость.
Соотношение между различными типами мышечных клеток у каждого человека генетически предопределено. Однако, используя физические нагрузки определенного характера, можно целенаправленно вызывать изменение спектра мышечных волокон. За счет применения силовых упражнений происходит смещение этого спектра в сторону преобладания белых волокон, имеющих больший диаметр по сравнению с красными и переходными, что в итоге приводит к гипертрофии тренируемых мышц. Основной причиной гипертрофии в этом случае является увеличение содержания в мышечных клетках сократительных элементов – миофибрилл. Поэтому мышечная гипертрофия, вызываемая силовыми нагрузками, относится к миофибриллярному типу.
Физические нагрузки, применяемые для развития мышечной гипертрофии миофибриллярного типа, на биохимическом уровне должны приводить к повреждению миофибрилл с последующей их суперкомпенсацией. С этой целью используются различные упражнения с отягощением.
Для развития силы часто используется метод повторных упражнений с напряжением 80–90% от максимальной силы. Наиболее эффективное отягощение – 85% от максимальной силы. В этом случае число повторений "до отказа" обычно 7–8. Каждое упражнение выполняется сериями, количество которых колеблется от 5 до 10, с интервалом отдыха между ними в несколько минут. Скорость выполнения упражнений определяется целью тренировки. Для преимущественного увеличения мышечной массы упражнения выполняются в медленном или умеренном темпе. Для одновременного развития силы и быстроты упражнения проводятся во взрывчато-плавном режиме: начальная фаза движения выполняется с большой скоростью, а завершается оно как можно более плавно. Поэтому в скоростно-силовых видах спортсмены в период силовой подготовки должны отказаться от медленного выполнения силовых упражнений, так как в этом случае утрачивается способность мышц к быстрому сокращению.
Время восстановления после скоростно-силовой тренировки составляет 2–3 дня. Однако, меняя мышечные группы, на которые направлены нагрузки, тренировочные занятия можно проводить через меньшие интервалы отдыха.
Обязательным условием эффективной силовой подготовки является полноценное, богатое белками питание, так как миофибриллы состоят исключительно из белков. Имеются данные о том, что развитию мышечной гипертрофии способствует ультрафиолетовое облучение. Предполагается, что под воздействием ультрафиолета увеличивается образование мужских половых гормонов, стимулирующих в организме синтез белков.
21 Биохимические основы быстроты (скорости) как качества двигательной деятельности
Быстрота как двигательное качество – это способность человека совершать двигательное действие в минимальный для данных условий отрезок времени с определенной частотой и импульсивностью. В вопросе о природе этого качества среди специалистов нет единства взглядов. Одни высказывают мысль, что физиологической основой быстроты является лабильность нервно-мышечного аппарата. Другие полагают, что важную роль в проявлении быстроты играет подвижность нервных процессов. Многочисленными исследованиями доказано, что быстрота является комплексным двигательным качеством человека.
Основные формы проявления быстроты человека – время двигательной реакции, время максимально быстрого выполнения одиночного движения, время выполнения движения с максимальной частотой, время выполнения целостного двигательного акта. Выделяют также еще одну форму проявления быстроты («скоростных качеств») – быстрое начало движения (то, что в спортивной практике называют «резкостью»). Практически наибольшее значение имеет скорость целостных двигательных актов (бег, плавание и др.), а не элементарные формы проявления быстроты, хотя скорость целостного движения лишь косвенно характеризует быстроту человека.
Скелетная мышца представляет собой сложную систему, преоб химическую энергию в механическую работу и тепло. Основными компонентами мышечного волокна являются белки: актин и миозин.
При совершении быстрых движений для сокращения мышц требуется большое количество энергии в единицу времени при дефиците кислорода, поэтому основную роль при этом играют анаэробные процессы гидролиза АТФ.
Гидролиз АТФ в АТФазном центре головки миозина сопро изменением конформации последней и переводом ее в новое, высокоэнергетическое состояние. Повторное присоединение миозиновой головки к новому центру на актиновом филаменте вновь приводит к вращению головки, которое обеспечивается за в ней энергией. В каждом цикле соединения и разъ головки миозина с актином расщепляется одна молекула АТФ на каждый мостик. Быстрота вращения определяется скоро расщепления АТФ. Очевидно, что быстрые фазические во потребляют значительно больше АТФ в единицу времени и сохраняют меньше химической энергии во время тонической нагрузки, чем медленные волокна. Таким образом, в процессе хемомеханического преобразования АТФ обеспечивает разъедине головки миозина и актинового филамента и энергетику для дальнейшего взаимодействия головки миозина с другим участком актинового филамента. Эти реакции возможны при концентрации кальция выше 106 моль/л.
Уровень развития быстроты, в конечном итоге, определяет успех в подавляющем большинстве видов спорта. Даже марафонец должен, возможно, быстрее пробежать свою дистанцию, сохраняя высокую «крейсерскую» скорость (под «крейсерской» скоростью подразумевается средняя скорость прохождения дистанции). И успех тяжелоатлета зависит от того, с какой скоростью он сумеет выполнить необходимое движение.
Быстрота определяется:
а) путем измерения скорости движения в ответ на определенный сигнал реакциометрами различной конструкции;
б) по количеству движений за установленное время незагруженной конечностью или туловищем в границах определенной амплитуды;
в) по времени преодоления установленного короткого расстояния (например, бега на 20, 30 м);
г) по скорости выполнения однократного движения в сложном действии,например отталкивания в прыжках, движения плечевого пояса и руки в метаниях, удара в боксе, начального движения бегуна на короткие дистанции, движений гимнаста и др.
Все проявления быстроты эффективно развиваются при игре в баскетбол. Можно также порекомендовать ручной мяч, настольный теннис, подвижные игры с быстро меняющейся игровой ситуацией и быстрым передвижением. Главная задача при воспитании быстроты состоит в том, чтобы спортсмен преждевременно не специализировался в каком-либо одном упражнении скоростного характера, чтобы не включать в большом объеме однотипное повторение этого упражнения. Поэтому столь важно, чтобы спортсмены применяли скоростные упражнения возможно чаще в форме состязания или игры. В программу занятий должны входить в значительном объеме такие скоростные упражнения, как спринтерский бег со старта и с хода, бег с ускорением, прыжки в длину и высоту с предельно быстрым отталкиванием, метание облегченных снарядов, подвижные и спортивные игры, предельно быстро выполняемые акробатические упражнения и разнообразные специальные подготовительные упражнения.
Важное значение для воспитания быстроты и повышения скорости движений имеет правильное определение дозировки скоростных упражнений. Те из них, которые выполняются с максимальной интенсивностью, являются сильно действующим средством, вызывающим быстрое утомление. Это же относится и к упражнениям, направленным на повышение скорости движений. Поэтому упражнения, выполняемые с максимальной скоростью, должны применяться часто, но в относительно небольшом объеме. Длительность интервалов отдыха обусловлена степенью возбудимости центральной нервной системы и восстановлением показателей вегетативных функций, связанных с ликвидацией кислородного дефицита. Тренировочную работу для развития быстроты следует заканчивать, как только субъективные ощущения спортсмена или показания секундомера скажут об уменьшении установленной или максимальной быстроты.
22.биохимические основы качества выносливости к длительным нагрузкам и пути его развития
Выносливость – важнейшее двигательное качество, от уровня развития которого во многом зависят достижения атлета. Выносливость можно определить как время работы с заданной мощностью до появления утомления.
В соответствии с характером выполняемой работы выделяют общую и специальную выносливость. Общая выносливость отражает способность спортсмена выполнять неспецифические нагрузки. Такими нагрузками, например, для футболиста могут быть кросс, лыжные гонки, плавание, подвижные игры и т.п., а также выполнение физической работы бытового характера. Специальная выносливость характеризует выполнение физических нагрузок, специфических для определенного вида спорта и требующих технической, тактической и психологической подготовки спортсмена.
Первостепенное значение для проявления выносливости имеет уровень развития молекулярных механизмов образования АТФ – непосредственного источника энергии для обеспечения мышечного сокращения и расслабления
В зависимости от способа энергообеспечения выполняемой работы выделяют алактатную, лактатную и аэробную выносливость. Нередко используются термины." алактатиый, лактатный и аэробный компоненты выносливости.
Алактатная выносливость характеризуется наибольшим временем работы в зоне максимальной мощности. В зависимости от вида нагрузки можно выделить скоростную, скорости о-силовую и силовую алактатную выносливость. Главным источником энергии при мышечной работе максимальной мощности является креатинфосфатная реакция. Поэтому развитие алактатной выносливости обусловлено внутримышечными запасами креатинфосфата. Как уже отмечалось, более богаты креатинфосфатом белые мышечные волокна. В связи с этим большей алактатной выносливостью обладают мышцы с преобладанием белых волокон. Содержание креатинфосфата в мышцах можно существенно повысить, используя специальные упражнения. Принцип построения такой тренировки в интервальном режиме был описан выше, при рассмотрении энергообеспечения скоростно-силовых качеств.
Биохимическая оценка алактатной выносливости может быть дана путем определения суточного выделения с мочой креатинина. Этот показатель характеризует общие запасы в организме креатинфосфата. Рост алактатной выносливости обычно сопровождается увеличением суточного выделения креатинина. Другим критерием, характеризующим развитие алактатной выносливости, является алактатный кислородный долг, измеренный после завершения работы максимальной мощности.
Лактатная выносливость характеризует выполнение физических нагрузок в зоне субмаксимальной мощности. Основным источником энергии при работе с такой мощностью служит анаэробный распад мышечного гликогена до молочной кислоты, называемый гликолизом. Возможности гликолитического способа получения АТФ в значительной степени зависят от запасов мышечного гликогена. Чем выше дорабочая концентрация гликогена в мышцах, тем дольше он будет использоваться в гликолизе. Отсюда следует, что мышцы с преобладанием белых, богатых креатинфосфатом и гликогеном волокон обладают также и выраженной лактатной выносливостью. Другим фактором, определяющим лактатную выносливость, является резистентность мышечных клеток и всего организма в целом к возрастанию кислотности вследствие накопления лактата в мышцах и в крови.
Исходя из такой зависимости тренировки, направленные на развитие лактатной выносливости, строятся так, чтобы обеспечить выполнение двух задач. Во-первых, за счет выполняемых физических нагрузок в мышцах должно увеличиваться содержание гликогена. Во-вторых, тренировочные занятия должны привести к возникновению резистентности к накоплению лактата и повышению кислотности.
С этой целью применяются упражнения, вызывающие, с одной стороны, значительное исчерпание запасов мышечного гликогена, что является необходимым условием для его последующей суперкомпенсации, а с другой – приводящие к образованию больших количеств молочной кислоты. Таковыми являются физические нагрузки субмаксимальной мощности, выполняемые в интервальном или повторном режиме. Тренировка такого типа описана выше, при рассмотрении энергообеспечения скоростно-силовых качеств. В зависимости от характера применяемых нагрузок можно преимущественно развивать силовой или скоростной компонент лактатной выносливости.
Ведущим биохимическим показателем проявления лактатной выносливости при работе является накопление лактата в крови. Определение концентрации молочной кислоты в крови проводят после выполнения физической работы субмаксимальной мощности "до отказа". Высокий уровень концентрации молочной кислоты в крови свидетельствует об использовании для получения энергии во время работы больших количеств мышечного гликогена и развитии резистентности к возрастанию кислотности.
Такую же информацию можно получить, определяя в крови после субмаксимальных нагрузок изменение кислотно-щелочного баланса. В этом случае высокой лактатной выносливости соответствует значительный сдвиг водородного показателя крови в кислую сторону. Еще одним показателем развития лактатной выносливости может служить лактатный кислородный долг, измеренный после выполнения работы субмаксимальной мощности "до отказа". Чем выше значение этого показателя, тем больше вклад анаэробного распада гликогена в энергообеспечение проделанной работы. У спортсменов с хорошей физической подготовкой величины лактатного кислородного долга могут достигать 18–20 л.
В спортивной практике очень часто алактатную и лактатную выносливость объединяют в анаэробную.
Аэробная выносливость проявляется при выполнении продолжительных упражнений умеренной мощности, которые главным образом обеспечиваются энергией за счет аэробного окисления. Вклад анаэробного энергообразования ограничивается лишь начальным периодом врабатывания. В спортивной литературе зачастую под термином "выносливость" подразумевается именно аэробная выносливость.
Аэробная выносливость определяется тремя главнейшими факторами: запасами в организме доступных источников энергии, доставкой кислорода в работающие мышцы и развитием в работающих мышцах митохондриального окисления.
В качестве источников энергии обычно используются углеводы, жирные кислоты, кетоновые тела и аминокислоты. Вследствие большой продолжительности аэробной работы эти энергетические субстраты доставляются в мышцы кровью, так как собственные энергетические ресурсы мышечных клеток расходуются в начале работы.
В обеспечении мышц источниками энергии существенная роль принадлежит печени. Именно здесь во время выполнения длительных нагрузок происходит распад гликогена до глюкозы, которая затем с током крови поступает в скелетные мышцы и другие органы, участвующие в обеспечении мышечной деятельности. Другой процесс, протекающий в печени во время работы, окисление жирных кислот, сопровождающееся образованием кетоновых тел, которые также являются важными источниками энергии. Кроме того, в печени во время работы протекают и другие химические процессы, способствующие выполнению мышечной работы. В связи с такой важной ролью печени в обеспечении физической работы в спортивной практике применяют гепатопротекторы – фармакологические средства, улучшающие функционирование печени и ускоряющие в ней процессы восстановления.
Доставка кислорода в мышцы осуществляется кардиореспираторной системой. Поэтому для проявления аэробной выносливости исключительно важное значение имеет функциональное состояние сердечнососудистой и дыхательной систем, кислородная емкость крови, обусловленная количеством эритроцитов и содержанием в них гемоглобина.
Развитие аэробной выносливости в значительной мере определяется также состоянием нервно-гормональной регуляции. Ведущую роль в этой регуляции выполняют надпочечники, выделяющие в кровь катехолстины и глюкокортикоиды – гормоны, вызывающие перестройку организма, направленную на создание оптимальных условий для мышечной деятельности. Для проявления аэробной выносливости важна способность надпочечников в течение длительного времени поддерживать в кровяном русле повышенную концентрацию этих гормонов.
Внутримышечными факторами, ответственными за аэробную выносливость, являются размер и количество митохондрий – внутриклеточных структур, в которых при участии кислорода происходит синтез АТФ, а также содержание миоглобина – мышечного белка, обеспечивающего внутри мышечных волокон перенос кислорода к митохондриям. Как уже отмечалось, более высоким содержанием митохондрий и миоглобина характеризуются красные мышечные волокна. Отсюда вытекает, что более высокая аэробная выносливость наблюдается в мышцах с преобладанием красных волокон.
Аэробная выносливость в отличие от анаэробной менее специфична. Это обусловлено тем, что ее в большой мере лимитируют различные внемышечные факторы: функциональное состояние кардиореспираторной системы, печени и нервно-гормональной регуляции, кислородная емкость крови, запасы в организме легкодоступных источников энергии. Поэтому спортсмен, имеющий хороший уровень аэробной выносливости, может проявить ее не только в том виде деятельности, где он прошел специализированную подготовку, но и в других видах аэробной работы. Например, квалифицированный футболист может показать хороший результат в беге на длинные дистанции.
Многофакторность аэробной выносливости требует применения комплекса разнообразных тренировочных средств, поскольку каждое конкретное занятие, вызывая достаточно разностороннее воздействие на организм, все же преимущественно совершенствует одну какую-либо сторону функциональных возможностей. В итоге, тренировки, направленные на развитие аэробной выносливости, должны обеспечить повышение работоспособности кардиореспираторной системы, способствовать увеличению количества эритроцитов в крови и содержанию в них гемоглобина, росту концентрации миоглобина в мышечных клетках, лучшему обеспечению работающих органов энергетическими субстратами.
С этой целью применяются различные варианты повторной и интервальной тренировки, а также непрерывная длительная работа равномерной или переменной мощности.
В качестве примера построения тренировочных занятий, направленных на развитие аэробной выносливости, можно привести так называемую циркуляторную интервальную тренировку. Этот метод заключается в чередовании кратковременных упражнений небольшой интенсивности и длительностью от 30 до 90 с с интервалами отдыха такой же продолжительности. Такая работа стимулирует аэробное энергообеспечение мышечной деятельности и приводит к улучшению показателей кардиореспираторной системы.
Для повышения содержания в мышцах миоглобина может быть использована миоглобиновая интервальная тренировка. Спортсменам предлагаются очень короткие нагрузки средней интенсивности, чередуемые с такими же короткими промежутками отдыха. Выполняемые кратковременные нагрузки в основном обеспечиваются кислородом, который депонирован в мышечных клетках в форме комплекса с миоглобином. Короткий отдых между упражнениями достаточен для восполнения запасов кислорода.
Для увеличения кислородной емкости крови, а также для повышения концентрации миоглобина хороший эффект дают тренировки в условиях среднегорья.
Особенностью развития аэробной выносливости является возможность использования неспецифических упражнений, и в первую очередь подвижных игр, что позволяет сделать тренировочный процесс разнообразным и интересным.
Важную информацию для оценки аэробной выносливости можно Получить путем определения содержания и соотношения в крови основных энергетических субстратов в ходе выполнения продолжительной работы. У нетренированных людей между содержанием в крови глюкозы и продуктов мобилизации жира существуют реципрокные отношения. Высокая концентрация глюкозы в крови препятствует мобилизации жира из депо. Поэтому у нетренированных людей повышение содержания в крови жирных кислот, глицерина и кетоновых тел наблюдается только на фоне снижения концентрации глюкозы. У спортсменов, хорошо тренированных в аэробном режиме, мощная мобилизация жира отмечается на фоне не только нормального, но и повышенного содержания глюкозы в крови. Повышенная утилизация жира и кетоновых тел позволяет организму не только сохранить углеводы печени и крови, но и замедлить расходование мышечного гликогена, снижение концентрации которого является одним из факторов развития утомления.
В заключение необходимо отметить, что все компоненты выносливости наряду с рассмотренными выше энергетическими и структурными факторами в значительной мере зависят от технической, тактической и психологической подготовки. Хорошая техническая подготовка, правильно избранная тактика позволяет спортсмену экономно и рационально использовать энергетические резервы и тем самым дольше сохранять работоспособность. За счет высокой мотивации, большой силы воли спортсмен может продолжать выполнение работы даже в условиях наступления в организме значительных биохимических и функциональных сдвигов.
Похожие статьи